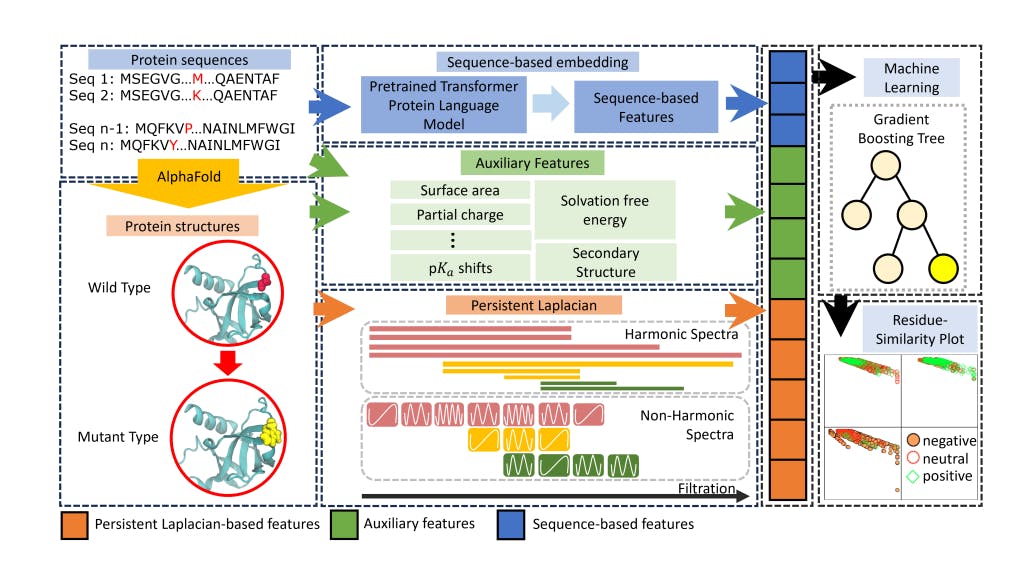
The Materials and Methods That We Used to Compare Tools for Fitting Mutational Signatures
by byMutation Technology Publications@mutation
byMutation Technology Publications@mutation
Mutation: process of changing in form or nature. We publish the best academic journals & first hand accounts of Mutation
March 18th, 2024






Mutation: process of changing in form or nature. We publish the best academic journals & first hand accounts of Mutation


Mutation: process of changing in form or nature. We publish the best academic journals & first hand accounts of Mutation
About Author

Mutation: process of changing in form or nature. We publish the best academic journals & first hand accounts of Mutation
Comments