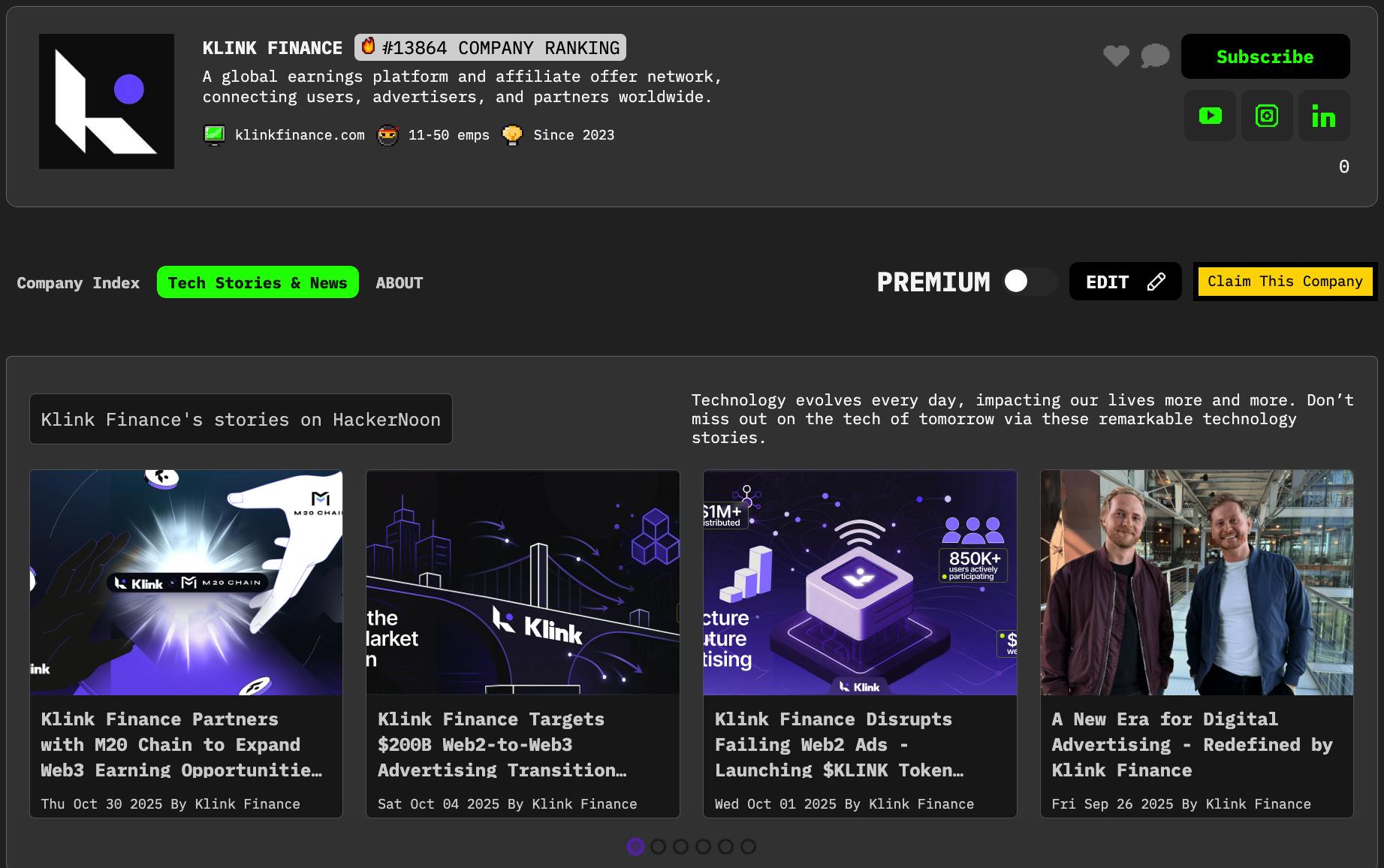
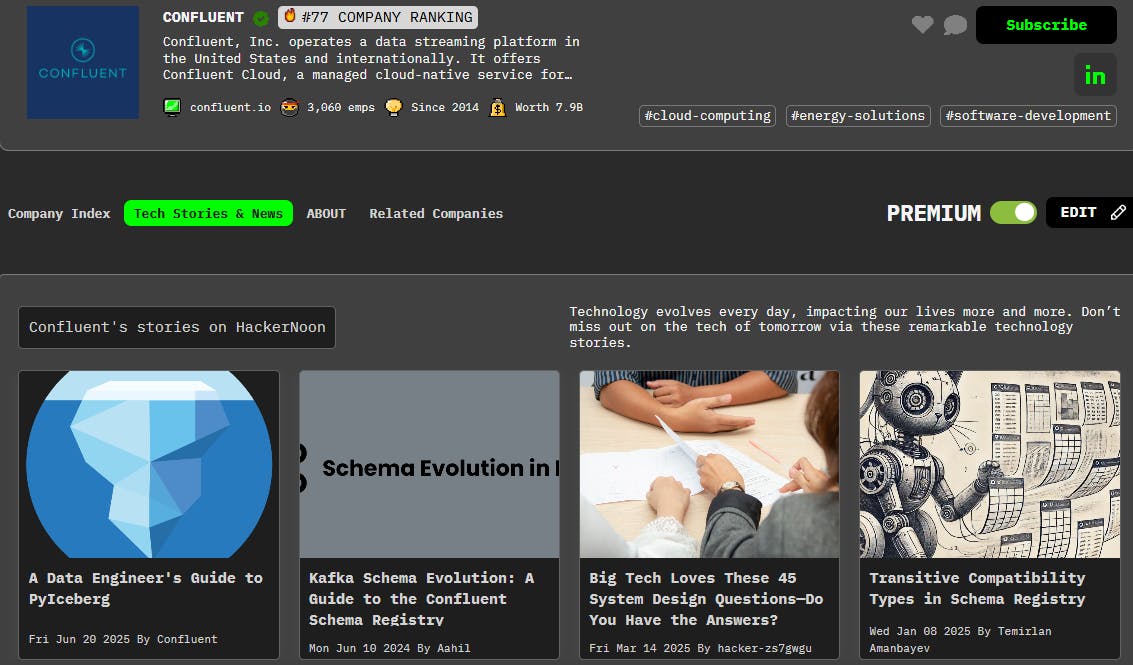
ผู้เขียน : มณฑลซานตง เฮน เลีย Ma ภาษาไทย J. J. Johannes Hjorth Alexander Kozlov ยุโรป He เซินเจิ้น Zhang เจนเน็ต Hellgren Kotaleski มณฑลซานตง สเตน Grillner เมื่อคุณ ภาษาไทย ผู้เขียน : มณฑลซานตง เฮน เลีย Ma ภาษาไทย จอห์นฮาโนส Hjorth อเล็กซานคอสโลว ยุโรป He เซินเจิ้น Zhang เจนเน็ต Hellgren Kotaleski มณฑลซานตง สเตน Grillner เมื่อคุณ ภาษาไทย สารสกัดจาก แบบจําลองหลายส่วนที่มีรายละเอียดทางชีวภาพเป็นเครื่องมือที่มีประสิทธิภาพในการสํารวจหลักการการคํานวณของสมองและยังทําหน้าที่เป็นกรอบทฤษฎีสําหรับการสร้างอัลกอริทึมสําหรับระบบอัจฉริยะเทียม (AI) อย่างไรก็ตามค่าใช้จ่ายการคํานวณที่แพง จํากัด การใช้งานทั้งในสาขาวิทยาศาสตร์ประสาทและสาขาเทียมอัจฉริยะ ข้อบกพร่องที่สําคัญในการจําลองแบบจําลองชิ้นส่วนรายละเอียดคือความสามารถของเครื่องจําลองในการแก้ปัญหาระบบขนาดใหญ่ของคํานวณเชิงเส้น ที่นี่เรานําเสนอเรื่องใหม่ แอนดริทิก ภูมิศาสตร์ วิธีการ cheduling (DHS) เพื่อเร่งกระบวนการดังกล่าวอย่างมีนัยสําคัญ เราพิสูจน์ทางทฤษฎีว่าการใช้งาน DHS เป็นการคํานวณที่เหมาะสมและแม่นยํา วิธีนี้ขึ้นอยู่กับ GPU ทํางานด้วยความเร็วที่สูงขึ้น 2-3 ครั้งกว่าวิธีการ Hines แบบคลาสสิกในแพลตฟอร์ม CPU แบบดั้งเดิม เราสร้างกรอบ DeepDendrite ซึ่งรวมวิธีการ DHS และเครื่องยนต์การคํานวณ GPU ของจําลอง NEURON และแสดงให้เห็นถึงการใช้งานของ DeepDendrite ในงานวิทยาศาสตร์ประสาท เราตรวจสอบว่ารูปแบบพื้นที่ของอินพุตกระดูกสันหลังมีผลต่อความกระตือรือร้นของประสาทประสาทในรูปแบบประสาทปิรามิดของมนุษย์ที่มีรายละเอียด 25,000 กระดูกสันหลัง นอกจากนี้เรายังให้คําพูดสั้นเกี่ยวกับศักยภาพของ D H S การแนะนํา การคํานวณหลักการการเข้ารหัสและคํานวณของประสาทประสาทเป็นสิ่งจําเป็นสําหรับวิทยาศาสตร์ประสาทสมองของสัตว์เลี้ยงประกอบด้วยมากกว่าพันชนิดของประสาทประสาทที่แตกต่างกันที่มีคุณสมบัติทางพยาธิวิทยาและทางชีวภาพที่ไม่ซ้ํากัน แม้ว่ามันจะไม่ถูกต้องในแนวคิดอีกต่อไป แต่หลักการ "ประสาทประสาทประสาท" ในฐานะที่เป็นหน่วยประสาทที่ได้รับการพิจารณาเป็นหน่วยรวมที่เรียบง่ายยังคงใช้กันอย่างแพร่หลายในการคํานวณประสาทโดยเฉพาะอย่างยิ่งในการวิเคราะห์เครือข่ายประสาท ในปีที่ผ่านมาอัจฉริยะเทียมที่ทันสมัย (AI) ได้ใช้หลักการนี้และพัฒนาเครื่องมือที่มีประสิทธิภาพเช่นเครือข่ายประสาทเทียม (ANN) อย่างไรก็ตามนอกเหนือจากการคํานวณที่ครอบคลุมในระดับประสาทเดียวห้องใต้เซลล์เช่น neuronal dendrites ยังสามารถดําเนินการกระบวนการที่ไม่ใช่เชิงเส้นเป็นหน่วยคํานวณอิสระ , , , , นอกจากนี้ spines dendritic, ออกขนาดเล็กที่หนาแน่นครอบคลุม dendrites ใน neurons spiny, สามารถ compartmentalize สัญญาณ synaptic ช่วยให้พวกเขาสามารถแยกออกจาก dendrites พ่อของพวกเขา ex vivo และ in vivo , , , . 1 2 3 4 5 6 7 8 9 10 11 Simulations using biologically detailed neurons provide a theoretical framework for linking biological details to computational principles. The core of the biophysically detailed multi-compartment model framework , ช่วยให้เราสามารถจําลอง neurons กับ morphologies dendritic จริง, การนําไอออนในร่ม, และ extrinsic อินพุต synaptic โครงกระดูกสันหลังของโมเดล multi-compartment รายละเอียด, นั่นคือ, dendrites, ก่อสร้างขึ้นบนคลาสสิก ทฤษฎีเคเบิล , ซึ่งจําลองคุณสมบัติ membrane biophysical ของ dendrites เป็นสายเคเบิลที่ผ่านการใช้งานให้คําอธิบายทางคณิตศาสตร์ของวิธีการสัญญาณอิเล็กทรอนิกส์โจมตีและแพร่กระจายผ่านกระบวนการประสาทที่ซับซ้อน โดยการรวมทฤษฎีสายเคเบิลกับกลไกทางชีวภาพที่ใช้งานเช่นช่องทางไอออนกระตุ้นและยับยั้งการไหลเวียน synaptic ฯลฯ แบบจําลองหลายส่วนรายละเอียดสามารถบรรลุการคํานวณประสาทเซลล์และเซลล์ใต้ภายนอกข้อ จํากัด ของการทดลอง , . 12 13 12 4 7 นอกเหนือจากผลกระทบที่รุนแรงของมันต่อวิทยาศาสตร์ประสาทรุ่นประสาทรายละเอียดทางชีวภาพเมื่อเร็ว ๆ นี้ถูกนํามาใช้เพื่อสะสมความแตกต่างระหว่างรายละเอียดโครงสร้างประสาทและประสาททางชีวภาพและ AI เทคนิคที่ครอบคลุมในสาขาอัจฉริยะอัจฉริยะที่ทันสมัยคือ ANNs ประกอบด้วยประสาทประสาทจุดซึ่งเป็นอะนาล็อกกับเครือข่ายประสาททางชีวภาพ แม้ว่า ANNs ที่มีอัลกอริทึม "backpropagation-of-error" (backprop) จะบรรลุประสิทธิภาพที่โดดเด่นในแอพพลิเคชันเฉพาะแม้จะเอาชนะผู้เล่นมืออาชีพชั้นนําของมนุษย์ในเกมของ Go และช็อต , สมองมนุษย์ยังคงเอาชนะ ANNs ในโดเมนที่เกี่ยวข้องกับสภาพแวดล้อมที่มีไดนามิกและเสียงรบกวนมากขึ้น , การศึกษาทางทฤษฎีล่าสุดแสดงให้เห็นว่าการบูรณาการ dendritic เป็นสิ่งสําคัญในการสร้างอัลกอริทึมการเรียนรู้ที่มีประสิทธิภาพซึ่งอาจเกิน backprop ในการประมวลผลข้อมูลแบบขนาน , , นอกจากนี้โมเดลหลายส่วนรายละเอียดเดียวสามารถเรียนรู้การคํานวณ nonlinear ในระดับเครือข่ายสําหรับประสาทประสาทจุดโดยการปรับความแข็งแรง synaptic เท่านั้น , ซึ่งแสดงให้เห็นถึงศักยภาพเต็มรูปแบบของโมเดลรายละเอียดในการสร้างระบบ AI ที่คล้ายกับสมองที่มีประสิทธิภาพมากขึ้น ดังนั้นจึงเป็นสิ่งสําคัญที่จะขยาย paradigms ใน AI ที่คล้ายกับสมองจากโมเดลประสาทรายละเอียดเดียวไปจนถึงเครือข่ายรายละเอียดทางชีวภาพขนาดใหญ่ 14 15 16 17 18 19 20 21 22 หนึ่งในความท้าทายที่ยาวนานของวิธีการจําลองรายละเอียดอยู่ในค่าใช้จ่ายการคํานวณที่สูงมากซึ่งได้ จํากัด การประยุกต์ใช้ของมันอย่างรุนแรงไปยังวิทยาศาสตร์ประสาทและ AI ข้อบกพร่องที่สําคัญของจําลองคือการแก้คํานวณเชิงเส้นบนพื้นฐานของทฤษฎีพื้นฐานของการจําลองรายละเอียด , , เพื่อปรับปรุงประสิทธิภาพวิธีการ Hines คลาสสิคช่วยลดความซับซ้อนเวลาในการแก้ปัญหาสม่ําเสมอจาก O(n3) ถึง O(n) ซึ่งได้รับการนํามาใช้กันอย่างแพร่หลายเป็นอัลกอริทึมหลักในเครื่องจําลองที่นิยมเช่น NEURON และ Genesis อย่างไรก็ตามวิธีการนี้ใช้วิธีการตามลําดับเพื่อประมวลผลแต่ละห้องตามลําดับ เมื่อการจําลองเกี่ยวข้องกับ dendrites หลายรายละเอียดทางชีวภาพที่มีสปิน dendritic ตารางการสัดส่วนเชิงเส้น (“Hines Matrix”) จะสแกนตามลําดับด้วยจํานวน dendrites หรือสปินที่เพิ่มขึ้น (ภาพ. ), ทําให้วิธีการ Hines ไม่ได้ใช้งานได้อีกต่อไปเนื่องจากมีภาระหนักมากต่อการจําลองทั้งหมด 12 23 24 25 26 1E แบบจําลอง neuron pyramidal ชั้น 5 และสูตรทางคณิตศาสตร์ที่ใช้กับแบบจําลอง neuron รายละเอียด กระแสการทํางานเมื่อจําลองแบบหมายเลขรุ่นประสาทรายละเอียด ขั้นตอนการแก้ปัญหาสม่ําเสมอเป็นขอบเขตในการจําลอง ตัวอย่างของสัดส่วนเชิงเส้นในการจําลอง ขึ้นอยู่กับข้อมูลของวิธีการ Hines เมื่อแก้ความสม่ําเสมอเชิงเส้น ขนาดของแม่เหล็ก Hines มีความซับซ้อนของโมเดล จํานวนของระบบสัดส่วนเชิงเส้นที่จะได้รับการแก้ปัญหามีการเพิ่มขึ้นอย่างมีนัยสําคัญเมื่อโมเดลจะเติบโตรายละเอียดมากขึ้น ค่าใช้จ่ายการคํานวณ (ขั้นตอนที่ดําเนินการในขั้นตอนการแก้ปัญหานี้) ของวิธีการ Hines ซีรี่ส์บนรูปแบบประสาทที่แตกต่างกัน Illustration of different solving methods. Different parts of a neuron are assigned to multiple processing units in parallel methods (mid, right), shown with different colors. In the serial method (left), all compartments are computed with one unit. ค่าใช้จ่ายการคํานวณของสามวิธีการ เมื่อแก้ความสม่ําเสมอของรูปแบบปิรามิดด้วยสกรู เวลาในการทํางานของวิธีการที่แตกต่างกันในการแก้ปัญหาสม่ําเสมอสําหรับ 500 แบบ Pyramidal กับ Spines เวลาในการทํางานแสดงให้เห็นการบริโภคเวลาของการจําลอง 1 วินาที (แก้สม่ําเสมอ 40,000 ครั้งด้วยขั้นตอนเวลาของ 0.025 มิลลิเมตร) วิธีการ parallel p-Hines ใน CoreNEURON (บน GPU), Branch based branch-based parallel method (บน GPU), DHS Dendritic method of scheduling (บน GPU) a b c d c e f g h g i ในช่วงทศวรรษที่ผ่านมาได้ประสบความสําเร็จอย่างมากในการเร่งวิธีการ Hines โดยใช้วิธีการที่สอดคล้องกันในระดับเซลล์ซึ่งช่วยให้การคํานวณของส่วนต่าง ๆ ในแต่ละเซลล์สามารถสอดคล้องกันได้ , , , , , อย่างไรก็ตามวิธีการขนานในระดับเซลล์ปัจจุบันมักจะขาดกลยุทธ์การขนานที่มีประสิทธิภาพหรือขาดความแม่นยําทางตัวเลขที่เพียงพอเมื่อเทียบกับวิธีการ Hines ที่เดิม 27 28 29 30 31 32 ที่นี่เราพัฒนาเครื่องมือจําลองแบบอัตโนมัติแบบเต็มรูปแบบที่แม่นยําและเพิ่มประสิทธิภาพซึ่งสามารถเร่งประสิทธิภาพการคํานวณอย่างมีนัยสําคัญและลดค่าใช้จ่ายในการคํานวณ นอกจากนี้เครื่องมือจําลองนี้สามารถนํามาใช้ได้อย่างราบรื่นเพื่อสร้างและทดสอบเครือข่ายประสาทที่มีรายละเอียดทางชีวภาพสําหรับการเรียนรู้เครื่องและแอปพลิเคชัน AI อย่างสําคัญเราจะกําหนดการคํานวณแบบขนานของวิธีการ Hines เป็นปัญหาการวางแผนทางคณิตศาสตร์และสร้างวิธีการ Dendritic Hierarchical Scheduling (DHS) ขึ้นอยู่กับการเพิ่มประสิทธิภาพการรวม และทฤษฎีการคํานวณแบบขนาน . เราแสดงให้เห็นว่าอัลกอริทึมของเราให้การวางแผนที่เหมาะสมโดยไม่ต้องสูญเสียความแม่นยําใด ๆ นอกจากนี้เรายังได้เพิ่มประสิทธิภาพ DHS สําหรับชิป GPU ที่ทันสมัยที่สุดในปัจจุบันโดยใช้เกียจหน่วยความจํา GPU และกลไกการเข้าถึงหน่วยความจํา รวมกัน DHS สามารถเร่งการคํานวณได้ 60-1,500 ครั้ง (ตารางเพิ่มเติม) ) เมื่อเทียบกับเครื่องจําลองคลาสสิก Neuron ในขณะที่รักษาความแม่นยําเดียวกัน 33 34 1 25 เพื่อให้สามารถใช้การจําลอง dendritic อย่างละเอียดเพื่อใช้ใน AI เราจะสร้างกรอบ DeepDendrite โดยรวมแพลตฟอร์ม CoreNEURON (เครื่องคอมพิวเตอร์ที่เพิ่มประสิทธิภาพสําหรับ NEURON) ที่บูรณาการ DHS เป็นเครื่องยนต์จําลองและสองโมดูลเสริม (โมดูล I / O และโมดูลการเรียนรู้) ที่สนับสนุนอัลกอริทึมการเรียนรู้ dendritic ในระหว่างการจําลอง DeepDendrite ทํางานบนแพลตฟอร์มฮาร์ดแวร์ GPU รองรับทั้งงานจําลองปกติในวิทยาศาสตร์ประสาทและงานเรียนรู้ใน AI 35 สุดท้าย แต่ไม่น้อยเรายังนําเสนอแอพพลิเคชันหลายอย่างโดยใช้ DeepDendrite ซึ่งมุ่งเน้นไปที่ความท้าทายที่สําคัญในวิทยาศาสตร์ประสาทและ AI: (1) เราแสดงให้เห็นว่ารูปแบบพื้นที่ของอินพุตกระดูกสันหลัง dendritic มีผลต่อกิจกรรมประสาทกับประสาทที่มีกระดูกสันหลังทั่วต้นไม้ dendritic (โมเดลกระดูกสันหลังเต็มรูปแบบ) DeepDendrite ช่วยให้เราสามารถสํารวจการคํานวณประสาทในการจําลองประสาทกระดูกสันหลังของมนุษย์ที่มี ~ 25,000 กระดูกสันหลัง dendritic (2) ในการสนทนาเรายังพิจารณาศักยภาพของ DeepDendrite ในแง่ของ AI โดยเฉพาะอย่างยิ่งในการสร้าง ANNs กับประสาทสันหลังของมนุษย์ที่มีรายละเอียดทางพยาธิวิทยา ผลลัพธ์ของเราแสดง รหัสแหล่งที่มาทั้งหมดสําหรับ DeepDendrite, รุ่นสมุนไพรเต็มรูปแบบและรุ่นเครือข่าย dendritic รายละเอียดสามารถใช้ได้โดยสาธารณะออนไลน์ (ดู Code Availability) กรอบการเรียนรู้แหล่งที่มาเปิดของเราสามารถบูรณาการได้อย่างง่ายดายกับกฎการเรียนรู้ dendritic อื่น ๆ เช่นกฎการเรียนรู้สําหรับ dendrites แบบไม่ตรง (เต็มรูปแบบ) , burst-dependent synaptic plasticity , and learning with spike prediction โดยรวมการศึกษาของเราให้ชุดเครื่องมือที่สมบูรณ์แบบที่มีศักยภาพที่จะเปลี่ยนระบบนิเวศวิทยาการคํานวณปัจจุบัน โดยใช้พลังงานของการคํานวณ GPU เราคาดหวังว่าเครื่องมือเหล่านี้จะช่วยให้การสํารวจหลักการการคํานวณในระดับระบบของโครงสร้างที่ละเอียดอ่อนของสมองเช่นเดียวกับการส่งเสริมการโต้ตอบระหว่างวิทยาศาสตร์ประสาทและเทียมอัจฉริยะสมัยใหม่ 21 20 36 ผล วิธี Dendritic Hierarchical Scheduling (DHS) การคํานวณของกระแสไอออนและการแก้ปัญหาของคํานวณเชิงเส้นเป็นสองขั้นตอนที่สําคัญเมื่อจําลอง neurons รายละเอียดทางชีวภาพซึ่งเป็นเวลานานและก่อให้เกิดภาระการคํานวณที่รุนแรง โชคดีที่การคํานวณของกระแสไอออนของแต่ละห้องเป็นกระบวนการที่เป็นอิสระอย่างสมบูรณ์เพื่อให้สามารถ parallelized โดยธรรมชาติบนอุปกรณ์ที่มีหน่วยการคํานวณแบบขนานขนาดใหญ่เช่น GPUs เป็นผลให้การแก้ปัญหาสม่ําเสมอจะกลายเป็นขั้วขวางที่เหลือสําหรับกระบวนการ parallelization (รูปภาพ) ) 37 1A-F เพื่อแก้ไขข้อบกพร่องนี้มีการพัฒนาวิธีการขนานในระดับเซลล์ซึ่งเร่งการคํานวณเซลล์เดี่ยวโดยการ "แบ่ง" เซลล์เดี่ยวเป็นหลายห้องที่สามารถคํานวณได้แบบขนาน , , อย่างไรก็ตามวิธีการดังกล่าวขึ้นอยู่กับความรู้ก่อนหน้านี้อย่างมากเพื่อสร้างกลยุทธ์การปฏิบัติเกี่ยวกับวิธีการแบ่ง neuron เดียวลงในห้อง (ภาพ. ; Fig พิเศษ ). ดังนั้นจึงกลายเป็นประสิทธิภาพน้อยกว่าสําหรับประสาทที่มีรูปร่างไม่สอดคล้องกันเช่นประสาท Pyramidal และประสาท Purkinje 27 28 38 1g−i 1 We aim to develop a more efficient and precise parallel method for the simulation of biologically detailed neural networks. First, we establish the criteria for the accuracy of a cellular-level parallel method. Based on the theories in parallel computing เรานําเสนอสามเงื่อนไขเพื่อให้แน่ใจว่าวิธีการที่คล้ายคลึงกันจะให้โซลูชั่นที่คล้ายคลึงกันกับวิธีการการคํานวณแบบซีรีส์ Hines ตามการขึ้นอยู่กับข้อมูลในวิธีการ Hines (ดู Methods) จากนั้นเพื่อประเมินเวลาทํางานทางทฤษฎีเช่นประสิทธิภาพของวิธีการการคํานวณแบบซีรีส์และแบบ parallel เราแนะนําและกําหนดแนวคิดของค่าใช้จ่ายการคํานวณเป็นจํานวนขั้นตอนที่วิธีการใช้ในการแก้คํานวณ (ดู Methods) 34 ขึ้นอยู่กับความแม่นยําของการจําลองและค่าใช้จ่ายการคํานวณเราจะ formulate the parallelization problem as a mathematical scheduling problem (see Methods). In simple terms, we view a single neuron as a tree with many nodes (divisions). สําหรับ ลวดขนานเราสามารถคํานวณได้สูงสุด nodes at each step, but we need to ensure a node is computed only if all its children nodes have been processed; our goal is to find a strategy with the minimum number of steps for the entire procedure. k k เพื่อสร้างพาร์ทิชันที่ดีที่สุดเราเสนอวิธีการที่เรียกว่า Dendritic Hierarchical Scheduling (DHS) (หลักฐานทางทฤษฎีจะนําเสนอใน Methods) ความคิดหลักของ DHS คือการให้ความสําคัญกับ nodes ลึก (ภาพ. วิธี DHS ประกอบด้วยสองขั้นตอน: การวิเคราะห์ topology dendritic และหาพาร์ทิชันที่ดีที่สุด: (1) ตามรูปแบบรายละเอียดเราจะได้รับต้นไม้ความสัมพันธ์ที่สอดคล้องกันและคํานวณความลึกของแต่ละแกน (ความลึกของแกนคือจํานวนแกนของต้นกําเนิดของมัน) บนต้นไม้ (ภาพ. ) (2) หลังจากวิเคราะห์ topology เราค้นหาผู้สมัครและเลือกมากที่สุด ปัจจัยการประมวลผลที่ลึกที่สุด (ปัจจัยการประมวลผลเป็นปัจจัยการประมวลผลเฉพาะเมื่อมีการประมวลผลปัจจัยการประมวลผลทั้งหมด) กระบวนการนี้จะทําซ้ําจนกว่าจะประมวลผลปัจจัยการประมวลผลทั้งหมด ) 2a 2B, C k 2d กระบวนการ DHS มุมผู้สมัครที่ลึกที่สุดของแต่ละ iteration ตัวอักษรของการคํานวณความลึกของ nodes ของรูปแบบที่แยกส่วน แบบแรกจะแปลงเป็นโครงสร้างต้นไม้แล้วความลึกของแต่ละ nodes จะคํานวณ สีแสดงค่าความลึกที่แตกต่างกัน Topology analysis on different neuron models. Six neurons with distinct morphologies are shown here. For each model, the soma is selected as the root of the tree so the depth of the node increases from the soma (0) to the distal dendrites. Illustration of performing DHS on the model in มีสี่เส้นด้าย ผู้สมัคร: หน่วยที่สามารถประมวลผลได้ ผู้สมัครที่เลือก: หน่วยที่เลือกโดย DHS นั่นคือ deepest candidates. Processed nodes: nodes that have been processed before. กลยุทธ์ paralelization ได้รับโดย DHS หลังจากกระบวนการใน DHS ลดขั้นตอนของการประมวลผลโซ่โซ่จาก 14 ถึง 5 โดยการกระจายโซ่ไปยังหลายโซ่ ค่าใช้จ่ายที่เกี่ยวข้องซึ่งหมายความว่าอัตราส่วนของค่าใช้จ่ายการคํานวณของ DHS กับอัตราส่วนของวิธีการ Hines ชุดเมื่อใช้ DHS กับจํานวนลวดที่แตกต่างกันในรูปแบบที่แตกต่างกัน a k b c d b k e d f ใช้แบบจําลองที่เรียบง่ายด้วย 15 หน่วยเป็นตัวอย่างโดยใช้วิธีการคํานวณซีรีส์ Hines จะใช้ 14 ขั้นตอนในการประมวลผล nodes ทั้งหมดในขณะที่ใช้ DHS ที่มีสี่หน่วยขนานสามารถแบ่ง nodes ของมันเป็นห้าชุดย่อย (ภาพ. ): {{9,10,12,14}, {1,7,11,13}, {2,3,4,8}, {6}, {5}} เนื่องจาก nodes ใน subset เดียวกันสามารถประมวลผลได้ใน paralel มันใช้เวลาเพียงห้าขั้นตอนในการประมวลผล nodes ทั้งหมดโดยใช้ DHS (ภาพ. ) 2d 2E Next, we apply the DHS method on six representative detailed neuron models (selected from ModelDB ) ด้วยจํานวนแถบที่แตกต่างกัน (ภาพ. ): รวมถึง neurons pyramidal cortical และ hippocampal , , , cerebellar Purkinje neurons เซลล์ประสาท Streathal Projection Neurons (SPN) ), and olfactory bulb mitral cells , ครอบคลุมประสาทหลักที่สําคัญในพื้นที่ความรู้สึก cortical และ subcortical จากนั้นเราวัดค่าใช้จ่ายการคํานวณ ค่าใช้จ่ายการคํานวณที่เกี่ยวข้องที่นี่ถูกกําหนดโดยอัตราส่วนของค่าใช้จ่ายการคํานวณของ DHS กับวิธีการ Hines ซีรี่ส์ ค่าใช้จ่ายการคํานวณเช่นจํานวนขั้นตอนที่ดําเนินการในการแก้ปัญหาคํานวณลดลงอย่างรุนแรงด้วยการเพิ่มจํานวนเส้นด้าย ตัวอย่างเช่นด้วยเส้นด้าย 16 ค่าใช้จ่ายการคํานวณของ DHS คือ 7%-10% เมื่อเทียบกับวิธีการ Hines ซีรี่ส์ ที่น่าสนใจวิธีการ DHS มาถึงขอบเขตต่ําของค่าใช้จ่ายการคํานวณของพวกเขาสําหรับประสาทที่นําเสนอเมื่อได้รับเส้นด้าย 16 หรือแม้แต่ 8 (ภาพ. ), suggesting adding more threads does not improve performance further because of the dependencies between compartments. 39 2f 40 41 42 43 44 45 2F ร่วมกันเราสร้างวิธีการ DHS ที่ช่วยให้การวิเคราะห์แบบอัตโนมัติของ topology dendritic และพาร์ทิชันที่ดีที่สุดสําหรับการคํานวณแบบขนาน เป็นมูลค่าสังเกตว่า DHS ค้นหาพาร์ทิชันที่เหมาะสมก่อนที่จะเริ่มต้นการจําลองและไม่จําเป็นต้องใช้การคํานวณเพิ่มเติมเพื่อแก้คํานวณ การเร่ง DHS โดยการเพิ่มหน่วยความจํา GPU DHS การคํานวณแต่ละประสาทด้วยเส้นด้ายหลายซึ่งใช้จํานวนมากของเส้นด้ายเมื่อเรียกใช้การจําลองเครือข่ายประสาท หน่วยประมวลผลกราฟิก (GPUs) ประกอบด้วยหน่วยประมวลผลขนาดใหญ่ (เช่นโปรเซสเซอร์สตรีมมิ่ง SPs, รูป ) สําหรับการคํานวณแบบขนาน . In theory, many SPs on the GPU should support efficient simulation for large-scale neural networks (Fig. อย่างไรก็ตามเราสังเกตเห็นอย่างต่อเนื่องว่าประสิทธิภาพของ DHS ลดลงอย่างมีนัยสําคัญเมื่อขนาดเครือข่ายเติบโตซึ่งอาจเป็นผลมาจากการจัดเก็บข้อมูลที่กระจายหรือการเข้าถึงหน่วยความจําเพิ่มเติมที่เกิดจากการโหลดและเขียนผลลัพธ์กลาง (ภาพ. ขวา ) 3A, B 46 3c 3d GPU architecture and its memory hierarchy. Each GPU contains massive processing units (stream processors). Different types of memory have different throughput. Architecture of Streaming Multiprocessors (SMs) Each SM contains multiple streaming processors, registers, and L1 cache. การประยุกต์ใช้ DHS บนสองประสาทแต่ละประสาทมีสี่เส้นด้าย ในระหว่างการจําลองแต่ละเส้นด้ายดําเนินการบนเครื่องประมวลผลสตรีม กลยุทธ์การเพิ่มประสิทธิภาพของหน่วยความจําบน GPU แผงด้านบนการกําหนดค่าเส้นด้ายและการจัดเก็บข้อมูลของ DHS ก่อน (ซ้าย) และหลัง (ขวา) การเพิ่มประสิทธิภาพของหน่วยความจํา ด้านล่างเป็นตัวอย่างของขั้นตอนเดียวใน triangularization เมื่อจําลองสองประสาทในระบบประสาท โปรเซสเซอร์ส่งคําขอข้อมูลเพื่อโหลดข้อมูลสําหรับแต่ละหัวข้อจากหน่วยความจําทั่วโลก โดยไม่ต้องเพิ่มหน่วยความจํา (ซ้าย) จะใช้เวลาเจ็ดธุรกรรมเพื่อโหลดข้อมูลการร้องขอทั้งหมดและบางธุรกรรมเพิ่มเติมสําหรับผลลัพธ์กลาง ด้วยการเพิ่มหน่วยความจํา (ขวา) จะใช้เวลาเพียงสองธุรกรรมเพื่อโหลดข้อมูลการร้องขอทั้งหมดบันทึกจะใช้สําหรับผลลัพธ์กลางซึ่งจะช่วยเพิ่มประสิทธิภาพของหน่วยความจํา เวลาทํางานของ DHS (32 เส้นต่อเซลล์แต่ละเซลล์) พร้อมและไม่มีหน่วยความจําเพิ่มขึ้นในหลายชั้น 5 รูปแบบปิรามิดที่มีสปิน ความเร็วเพิ่มขึ้นของหน่วยความจําเพิ่มขึ้นในหลายชั้น 5 รูปแบบปิรามิดที่มีหมุน การเพิ่มขึ้นของหน่วยความจํานําความเร็วเพิ่มขึ้น 1.6-2 ครั้ง a b c d d e f เราแก้ปัญหานี้โดยการเพิ่มหน่วยความจํา GPU, วิธีที่จะเพิ่มการไหลของหน่วยความจําโดยใช้เกียร์หน่วยความจําของ GPU และกลไกการเข้าถึง ขึ้นอยู่กับกลไกการโหลดหน่วยความจําของ GPU ลวดลําดับการโหลดข้อมูลที่สอดคล้องกันและจัดเก็บตามลําดับนําไปสู่การไหลของหน่วยความจําสูงเมื่อเทียบกับการเข้าถึงข้อมูลที่จัดเก็บโดยกระจายซึ่งช่วยลดการไหลของหน่วยความจํา , เพื่อให้บรรลุการส่งผ่านสูงเราจะจัดลําดับคําสั่งการคํานวณของ nodes และจัดลําดับใหม่ตามจํานวน nodes บนพวกเขา จากนั้นเราจะส่งผ่านการจัดเก็บข้อมูลในหน่วยความจําระดับโลกตามคําสั่งการคํานวณซึ่งหมายความว่า nodes ที่ได้รับการประมวลผลในขั้นตอนเดียวกันจะถูกจัดเก็บตามลําดับในหน่วยความจําระดับโลก นอกจากนี้เรายังใช้บันทึก GPU เพื่อจัดเก็บผลลัพธ์กลางเพื่อเสริมสร้างการส่งผ่านหน่วยความจําเพิ่มเติม ตัวอย่างนี้แสดงให้เห็นว่าการเพิ่มหน่วยความจําใช้เวลาเพียงสองธุรกรรมหน่วยความจําเพื่อโหลดข้อมูลคําขอแปด (ภาพ. นอกจากนี้การทดลองเกี่ยวกับตัวเลขหลายของประสาท Pyramidal กับ spin และรูปแบบประสาททั่วไป (ภาพ. ; Fig พิเศษ ) แสดงให้เห็นว่าการเพิ่มหน่วยความจําบรรลุความเร็ว 1.2-3.8 เท่าเมื่อเทียบกับ DHS ที่สุภาพ 46 47 3d 3E, F 2 เพื่อทดสอบประสิทธิภาพของ DHS ด้วยการเพิ่มหน่วยความจํา GPU เราเลือก 6 รุ่นประสาททั่วไปและประเมินเวลาทํางานของการแก้ปัญหาสัดส่วนสายเคเบิลในตัวเลขขนาดใหญ่ของแต่ละรุ่น ). เราตรวจสอบ DHS กับสี่เส้นด้าย (DHS-4) และสิบหกเส้นด้าย (DHS-16) สําหรับแต่ละประสาทตามลําดับ เมื่อเทียบกับวิธีการ GPU ใน CoreNEURON, DHS-4 และ DHS-16 สามารถเร่งความเร็วประมาณ 5 และ 15 ครั้งตามลําดับ (ภาพ. นอกจากนี้เมื่อเปรียบเทียบกับวิธีการ Hines ซีรี่ส์แบบดั้งเดิมใน NEURON ที่ทํางานกับ CPU แบบเดี่ยว DHS จะเพิ่มความเร็วในการจําลองด้วย 2-3 ช่วงขนาด (ตัวอักษรเพิ่มเติม) ), ในขณะที่รักษาความแม่นยําตัวเลขที่คล้ายกันในกรณีที่มีสกรูหนาแน่น (ตัวเลขเสริม. and ), active dendrites (Supplementary Fig. ) และกลยุทธ์การแบ่งแยกต่างหาก (เพิ่มเติม Fig. ) 4 4a 3 4 8 7 7 Run time of solving equations for a 1 s simulation on GPU (dt = 0.025 ms, 40,000 iterations in total). CoreNEURON: the parallel method used in CoreNEURON; DHS-4: DHS with four threads for each neuron; DHS-16: DHS with 16 threads for each neuron. , การแสดงภาพการแยกโดย DHS-4 และ DHS-16 แต่ละสีแสดงให้เห็นถึงเส้นด้ายเดียว ในระหว่างการคํานวณแต่ละเส้นด้ายสลับระหว่างสาขาที่แตกต่างกัน a b c DHS สร้างการแบ่งส่วนที่เหมาะสมโดยเฉพาะประเภทเซลล์ To gain insights into the working mechanism of the DHS method, we visualized the partitioning process by mapping compartments to each thread (every color presents a single thread in Fig. ). การแสดงภาพแสดงให้เห็นว่าลวดเดียวมักจะสลับระหว่างสาขาที่แตกต่างกัน (ภาพ. เป็นที่น่าตื่นตาตื่นใจที่ DHS สร้างส่วนแบ่งที่ตรงกันข้ามในประสาทประสาทแบบ symmetric เช่นประสาทประสาท streathal projection neuron (SPN) และเซลล์ Mitral (ภาพ. ). ในทางตรงกันข้าม, มันสร้างส่วนแบ่งของ neurons morphologically asymmetric เช่น neurons pyramidal และเซลล์ Purkinje (ภาพ. ), แสดงให้เห็นว่า DHS splits ต้นไม้ประสาทในระดับส่วนบุคคล (เช่นโหนดต้นไม้) แทนเป็นระดับสาขา ชิ้นส่วนที่เฉพาะเจาะจงในชนิดเซลล์นี้ช่วยให้ DHS สามารถใช้ประโยชน์อย่างเต็มที่ของเส้นด้ายที่มีอยู่ทั้งหมด 4B, C 4B, C 4B, C 4B, C โดยสรุปแล้ว DHS และ memory boosting สร้างโซลูชั่นที่ดีที่สุดที่พิสูจน์แล้วในทางทฤษฎีสําหรับการแก้ปัญหาสัดส่วนเชิงเส้นพร้อมกับประสิทธิภาพที่ไม่มีที่เห็นได้เปรียบ เมื่อใช้หลักการนี้เราได้สร้างแพลตฟอร์ม DeepDendrite ที่สามารถเข้าถึงได้แบบเปิดซึ่งสามารถใช้โดยนักวิทยาศาสตร์ประสาทในการใช้งานโมเดลโดยไม่มีความรู้เกี่ยวกับการเขียนโปรแกรม GPU ที่เฉพาะเจาะจง ด้านล่างเราแสดงให้เห็นถึงวิธีที่เราสามารถใช้ DeepDendrite ในงานประสาทวิทยาศาสตร์ นอกจากนี้เรายังพูดคุยเกี่ยวกับศักยภาพของกรอบ DeepDendrite สําหรับงานที่เกี่ยวข้องกับ AI ในส่วนการสนทนา DHS ช่วยให้การสร้างแบบจําลองระดับกระดูกสันหลัง As dendritic spines receive most of the excitatory input to cortical and hippocampal pyramidal neurons, striatal projection neurons, etc., their morphologies and plasticity are crucial for regulating neuronal excitability , , , , . However, spines are too small ( ~ 1 μm length) to be directly measured experimentally with regard to voltage-dependent processes. Thus, theoretical work is critical for the full understanding of the spine computations. 10 48 49 50 51 We can model a single spine with two compartments: the spine head where synapses are located and the spine neck that links the spine head to dendrites ทฤษฎีคาดการณ์ว่าคอกระดูกสันหลังที่บางมาก (0.1-0.5 มม. ในเส้นผ่าศูนย์กลาง) อิเล็กทรอนิกส์แยกหัวกระดูกสันหลังจาก dendrite พ่อแม่ของมันจึงแยกสัญญาณที่สร้างขึ้นที่หัวกระดูกสันหลัง . However, the detailed model with fully distributed spines on dendrites (“full-spine model”) is computationally very expensive. A common compromising solution is to modify the capacitance and resistance of the membrane by a ปัจจัย Spine , แทนที่จะจําลองสปริงทั้งหมดอย่างชัดเจน ที่นี่ spine factor aims at approximating the spine effect on the biophysical properties of the cell membrane . 52 53 F 54 F 54 Inspired by the previous work of Eyal et al. , we investigated how different spatial patterns of excitatory inputs formed on dendritic spines shape neuronal activities in a human pyramidal neuron model with explicitly modeled spines (Fig. ). Noticeably, Eyal et al. employed the ปัจจัยกระดูกสันหลังเพื่อรวมกระดูกสันหลังลงใน dendrites ในขณะที่เพียงไม่กี่กระดูกสันหลังที่เปิดใช้งานถูกเชื่อมโยงอย่างชัดเจนกับ dendrites (“รุ่นกระดูกสันหลังไม่กี่” ใน Fig. ). The value of spine in their model was computed from the dendritic area and spine area in the reconstructed data. Accordingly, we calculated the spine density from their reconstructed data to make our full-spine model more consistent with Eyal’s few-spine model. With the spine density set to 1.3 μm-1, the pyramidal neuron model contained about 25,000 spines without altering the model’s original morphological and biophysical properties. Further, we repeated the previous experiment protocols with both full-spine and few-spine models. We use the same synaptic input as in Eyal’s work but attach extra background noise to each sample. By comparing the somatic traces (Fig. ) and spike probability (Fig. ) in full-spine and few-spine models, we found that the full-spine model is much leakier than the few-spine model. In addition, the spike probability triggered by the activation of clustered spines appeared to be more nonlinear in the full-spine model (the solid blue line in Fig. ) than in the few-spine model (the dashed blue line in Fig. ). These results indicate that the conventional F-factor method may underestimate the impact of dense spine on the computations of dendritic excitability and nonlinearity. 51 5a F 5a F 5b, c 5d 5d 5d Experiment setup. We examine two major types of models: few-spine models and full-spine models. Few-spine models (two on the left) are the models that incorporated spine area globally into dendrites and only attach individual spines together with activated synapses. In full-spine models (two on the right), all spines are explicitly attached over whole dendrites. We explore the effects of clustered and randomly distributed synaptic inputs on the few-spine models and the full-spine models, respectively. Somatic voltages recorded for cases in . Colors of the voltage curves correspond to , scale bar: 20 ms, 20 mV. Color-coded voltages during the simulation in at specific times. Colors indicate the magnitude of voltage. Somatic spike probability as a function of the number of simultaneously activated synapses (as in Eyal et al.’s work) for four cases in . Background noise is attached. ระยะเวลาของการทดลองใน with different simulation methods. NEURON: conventional NEURON simulator running on a single CPU core. CoreNEURON: CoreNEURON simulator on a single GPU. DeepDendrite: DeepDendrite on a single GPU. a b a a c b d a e d In the DeepDendrite platform, both full-spine and few-spine models achieved 8 times speedup compared to CoreNEURON on the GPU platform and 100 times speedup compared to serial NEURON on the CPU platform (Fig. ; Supplementary Table ) while keeping the identical simulation results (Supplementary Figs. and ). Therefore, the DHS method enables explorations of dendritic excitability under more realistic anatomic conditions. 5e 1 4 8 Discussion In this work, we propose the DHS method to parallelize the computation of Hines method และเราแสดงให้เห็นว่า DHS ให้โซลูชั่นที่ดีที่สุดโดยไม่มีการสูญเสียความแม่นยําใด ๆ ต่อไปเราจะใช้ DHS บนแพลตฟอร์มฮาร์ดแวร์ GPU และใช้เทคนิคการเพิ่มหน่วยความจํา GPU เพื่อปรับปรุง DHS (รูปที่ 3 ) ). When simulating a large number of neurons with complex morphologies, DHS with memory boosting achieves a 15-fold speedup (Supplementary Table ) as compared to the GPU method used in CoreNEURON and up to 1,500-fold speedup compared to serial Hines method in the CPU platform (Fig. ; Supplementary Fig. and Supplementary Table ). Furthermore, we develop the GPU-based DeepDendrite framework by integrating DHS into CoreNEURON. Finally, as a demonstration of the capacity of DeepDendrite, we present a representative application: examine spine computations in a detailed pyramidal neuron model with 25,000 spines. Further in this section, we elaborate on how we have expanded the DeepDendrite framework to enable efficient training of biophysically detailed neural networks. To explore the hypothesis that dendrites improve robustness against adversarial attacks , we train our network on typical image classification tasks. We show that DeepDendrite can support both neuroscience simulations and AI-related detailed neural network tasks with unprecedented speed, therefore significantly promoting detailed neuroscience simulations and potentially for future AI explorations. 55 3 1 4 3 1 56 Decades of efforts have been invested in speeding up the Hines method with parallel methods. Early work mainly focuses on network-level parallelization. In network simulations, each cell independently solves its corresponding linear equations with the Hines method. Network-level parallel methods distribute a network on multiple threads and parallelize the computation of each cell group with each thread , . With network-level methods, we can simulate detailed networks on clusters or supercomputers . In recent years, GPU has been used for detailed network simulation. Because the GPU contains massive computing units, one thread is usually assigned one cell rather than a cell group , , . With further optimization, GPU-based methods achieve much higher efficiency in network simulation. However, the computation inside the cells is still serial in network-level methods, so they still cannot deal with the problem when the “Hines matrix” of each cell scales large. 57 58 59 35 60 61 Cellular-level parallel methods further parallelize the computation inside each cell. The main idea of cellular-level parallel methods is to split each cell into several sub-blocks and parallelize the computation of those sub-blocks , . However, typical cellular-level methods (e.g., the “multi-split” method ) pay less attention to the parallelization strategy. The lack of a fine parallelization strategy results in unsatisfactory performance. To achieve higher efficiency, some studies try to obtain finer-grained parallelization by introducing extra computation operations , , or making approximations on some crucial compartments, while solving linear equations , . These finer-grained parallelization strategies can get higher efficiency but lack sufficient numerical accuracy as in the original Hines method. 27 28 28 29 38 62 63 64 Unlike previous methods, DHS adopts the finest-grained parallelization strategy, i.e., compartment-level parallelization. By modeling the problem of “how to parallelize” as a combinatorial optimization problem, DHS provides an optimal compartment-level parallelization strategy. Moreover, DHS does not introduce any extra operation or value approximation, so it achieves the lowest computational cost and retains sufficient numerical accuracy as in the original Hines method at the same time. Dendritic spines are the most abundant microstructures in the brain for projection neurons in the cortex, hippocampus, cerebellum, and basal ganglia. As spines receive most of the excitatory inputs in the central nervous system, electrical signals generated by spines are the main driving force for large-scale neuronal activities in the forebrain and cerebellum , . The structure of the spine, with an enlarged spine head and a very thin spine neck—leads to surprisingly high input impedance at the spine head, which could be up to 500 MΩ, combining experimental data and the detailed compartment modeling approach , . Due to such high input impedance, a single synaptic input can evoke a “gigantic” EPSP ( ~ 20 mV) at the spine-head level , , thereby boosting NMDA currents and ion channel currents in the spine . However, in the classic single detailed compartment models, all spines are replaced by the coefficient modifying the dendritic cable geometries . This approach may compensate for the leak currents and capacitance currents for spines. Still, it cannot reproduce the high input impedance at the spine head, which may weaken excitatory synaptic inputs, particularly NMDA currents, thereby reducing the nonlinearity in the neuron’s input-output curve. Our modeling results are in line with this interpretation. 10 11 48 65 48 66 11 F 54 On the other hand, the spine’s electrical compartmentalization is always accompanied by the biochemical compartmentalization , , , resulting in a drastic increase of internal [Ca2+], within the spine and a cascade of molecular processes involving synaptic plasticity of importance for learning and memory. Intriguingly, the biochemical process triggered by learning, in turn, remodels the spine’s morphology, enlarging (or shrinking) the spine head, or elongating (or shortening) the spine neck, which significantly alters the spine’s electrical capacity , , , . Such experience-dependent changes in spine morphology also referred to as “structural plasticity”, have been widely observed in the visual cortex , โซมาโตเซนเซอร์ Cortex , , motor cortex , hippocampus , and the basal ganglia in vivo. They play a critical role in motor and spatial learning as well as memory formation. However, due to the computational costs, nearly all detailed network models exploit the “F-factor” approach to replace actual spines, and are thus unable to explore the spine functions at the system level. By taking advantage of our framework and the GPU platform, we can run a few thousand detailed neurons models, each with tens of thousands of spines on a single GPU, while maintaining ~100 times faster than the traditional serial method on a single CPU (Fig. ). Therefore, it enables us to explore of structural plasticity in large-scale circuit models across diverse brain regions. 8 52 67 67 68 69 70 71 72 73 74 75 9 76 5E Another critical issue is how to link dendrites to brain functions at the systems/network level. It has been well established that dendrites can perform comprehensive computations on synaptic inputs due to enriched ion channels and local biophysical membrane properties , , . For example, cortical pyramidal neurons can carry out sublinear synaptic integration at the proximal dendrite but progressively shift to supralinear integration at the distal dendrite นอกจากนี้ dendrites distal สามารถผลิตเหตุการณ์การฟื้นฟูเช่น dendritic sodium spikes, calcium spikes และ NMDA spikes / plateau potentials , . Such dendritic events are widely observed in mice or even human cortical neurons in vitro, which may offer various logical operations , or gating functions , . Recently, in vivo recordings in awake or behaving mice provide strong evidence that dendritic spikes/plateau potentials are crucial for orientation selectivity in the visual cortex , sensory-motor integration in the whisker system , , and spatial navigation in the hippocampal CA1 region . 5 6 7 77 6 78 6 79 6 79 80 81 82 83 84 85 To establish the causal link between dendrites and animal (including human) patterns of behavior, large-scale biophysically detailed neural circuit models are a powerful computational tool to realize this mission. However, running a large-scale detailed circuit model of 10,000-100,000 neurons generally requires the computing power of supercomputers. It is even more challenging to optimize such models for in vivo data, as it needs iterative simulations of the models. The DeepDendrite framework can directly support many state-of-the-art large-scale circuit models , , , which were initially developed based on NEURON. Moreover, using our framework, a single GPU card such as Tesla A100 could easily support the operation of detailed circuit models of up to 10,000 neurons, thereby providing carbon-efficient and affordable plans for ordinary labs to develop and optimize their own large-scale detailed models. 86 87 88 Recent works on unraveling the dendritic roles in task-specific learning have achieved remarkable results in two directions, i.e., solving challenging tasks such as image classification dataset ImageNet with simplified dendritic networks , and exploring full learning potentials on more realistic neuron , . However, there lies a trade-off between model size and biological detail, as the increase in network scale is often sacrificed for neuron-level complexity , , . Moreover, more detailed neuron models are less mathematically tractable and computationally expensive . 20 21 22 19 20 89 21 There has also been progress in the role of active dendrites in ANNs for computer vision tasks. Iyer et al. . proposed a novel ANN architecture with active dendrites, demonstrating competitive results in multi-task and continual learning. Jones and Kording used a binary tree to approximate dendrite branching and provided valuable insights into the influence of tree structure on single neurons’ computational capacity. Bird et al. . proposed a dendritic normalization rule based on biophysical behavior, offering an interesting perspective on the contribution of dendritic arbor structure to computation. While these studies offer valuable insights, they primarily rely on abstractions derived from spatially extended neurons, and do not fully exploit the detailed biological properties and spatial information of dendrites. Further investigation is needed to unveil the potential of leveraging more realistic neuron models for understanding the shared mechanisms underlying brain computation and deep learning. 90 91 92 In response to these challenges, we developed DeepDendrite, a tool that uses the Dendritic Hierarchical Scheduling (DHS) method to significantly reduce computational costs and incorporates an I/O module and a learning module to handle large datasets. With DeepDendrite, we successfully implemented a three-layer hybrid neural network, the Human Pyramidal Cell Network (HPC-Net) (Fig. ). This network demonstrated efficient training capabilities in image classification tasks, achieving approximately 25 times speedup compared to training on a traditional CPU-based platform (Fig. ; Supplementary Table ). 6a, b 6f 1 The illustration of the Human Pyramidal Cell Network (HPC-Net) for image classification. Images are transformed to spike trains and fed into the network model. Learning is triggered by error signals propagated from soma to dendrites. Training with mini-batch. Multiple networks are simulated simultaneously with different images as inputs. The total weight updates ΔW are computed as the average of ΔWi from each network. Comparison of the HPC-Net before and after training. Left, the visualization of hidden neuron responses to a specific input before (top) and after (bottom) training. Right, hidden layer weights (from input to hidden layer) distribution before (top) and after (bottom) training. Workflow of the transfer adversarial attack experiment. We first generate adversarial samples of the test set on a 20-layer ResNet. Then use these adversarial samples (noisy images) to test the classification accuracy of models trained with clean images. Prediction accuracy of each model on adversarial samples after training 30 epochs on MNIST (left) and Fashion-MNIST (right) datasets. Run time of training and testing for the HPC-Net. The batch size is set to 16. Left, run time of training one epoch. Right, run time of testing. Parallel NEURON + Python: training and testing on a single CPU with multiple cores, using 40-process-parallel NEURON to simulate the HPC-Net and extra Python code to support mini-batch training. DeepDendrite: training and testing the HPC-Net on a single GPU with DeepDendrite. a b c d e f Additionally, it is widely recognized that the performance of Artificial Neural Networks (ANNs) can be undermined by adversarial attacks —intentionally engineered perturbations devised to mislead ANNs. Intriguingly, an existing hypothesis suggests that dendrites and synapses may innately defend against such attacks . Our experimental results utilizing HPC-Net lend support to this hypothesis, as we observed that networks endowed with detailed dendritic structures demonstrated some increased resilience to transfer adversarial attacks compared to standard ANNs, as evident in MNIST and Fashion-MNIST datasets (Fig. ). This evidence implies that the inherent biophysical properties of dendrites could be pivotal in augmenting the robustness of ANNs against adversarial interference. Nonetheless, it is essential to conduct further studies to validate these findings using more challenging datasets such as ImageNet . 93 56 94 95 96 6d, e 97 In conclusion, DeepDendrite has shown remarkable potential in image classification tasks, opening up a world of exciting future directions and possibilities. To further advance DeepDendrite and the application of biologically detailed dendritic models in AI tasks, we may focus on developing multi-GPU systems and exploring applications in other domains, such as Natural Language Processing (NLP), where dendritic filtering properties align well with the inherently noisy and ambiguous nature of human language. Challenges include testing scalability in larger-scale problems, understanding performance across various tasks and domains, and addressing the computational complexity introduced by novel biological principles, such as active dendrites. By overcoming these limitations, we can further advance the understanding and capabilities of biophysically detailed dendritic neural networks, potentially uncovering new advantages, enhancing their robustness against adversarial attacks and noisy inputs, and ultimately bridging the gap between neuroscience and modern AI. Methods Simulation with DHS CoreNEURON simulator ( ) uses the NEURON เราใช้วิธีการ Dendritic Hierarchical Scheduling (DHS) ของเราในสภาพแวดล้อม CoreNEURON โดยการแก้ไขรหัสแหล่งของมัน รูปแบบทั้งหมดที่สามารถจําลองได้บน GPU ด้วย CoreNEURON สามารถจําลองได้ด้วย DHS โดยใช้คําสั่งต่อไปนี้: 35 https://github.com/BlueBrain/CoreNeuron 25 coreneuron_exec -d /path/to/models -e time --cell-permute 3 --cell-nthread 16 --gpu The usage options are as in Table . 1 Accuracy of the simulation using cellular-level parallel computation To ensure the accuracy of the simulation, we first need to define the correctness of a cellular-level parallel algorithm to judge whether it will generate identical solutions compared with the proven correct serial methods, like the Hines method used in the NEURON simulation platform. Based on the theories in parallel computing , a parallel algorithm will yield an identical result as its corresponding serial algorithm, if and only if the data process order in the parallel algorithm is consistent with data dependency in the serial method. The Hines method has two symmetrical phases: triangularization and back-substitution. By analyzing the serial computing Hines method , we find that its data dependency can be formulated as a tree structure, where the nodes on the tree represent the compartments of the detailed neuron model. In the triangularization process, the value of each node depends on its children nodes. In contrast, during the back-substitution process, the value of each node is dependent on its parent node (Fig. ). Thus, we can compute nodes on different branches in parallel as their values are not dependent. 34 55 1d Based on the data dependency of the serial computing Hines method, we propose three conditions to make sure a parallel method will yield identical solutions as the serial computing Hines method: (1) The tree morphology and initial values of all nodes are identical to those in the serial computing Hines method; (2) In the triangularization phase, a node can be processed if and only if all its children nodes are already processed; (3) In the back-substitution phase, a node can be processed only if its parent node is already processed. Once a parallel computing method satisfies these three conditions, it will produce identical solutions as the serial computing method. Computational cost of cellular-level parallel computing method To theoretically evaluate the run time, i.e., efficiency, of the serial and parallel computing methods, we introduce and formulate the concept of computational cost as follows: given a tree และ threads (basic computational units) to perform triangularization, parallel triangularization equals to divide the node set of into subsets, i.e., = { { , , … } where the size of each subset | | ≤ , i.e., at most nodes can be processed each step since there are only threads. The process of the triangularization phase follows the order: → → → → → , and nodes in the same subset can be processed in parallel. So, we define | | (the size of set , i.e., here) as the computational cost of the parallel computing method. In short, we define the computational cost of a parallel method as the number of steps it takes in the triangularization phase. Because the back-substitution is symmetrical with triangularization, the total cost of the entire solving equation phase is twice that of the triangularization phase. T k V T n V V1 V2 Vn Vi k k k V1 V2 Vn Vi V V n Mathematical scheduling problem ขึ้นอยู่กับความแม่นยําของการจําลองและค่าใช้จ่ายการคํานวณเราจะ formulate ปัญหา paralelization เป็น ปัญหาการกําหนดเวลาทางคณิตศาสตร์: Given a tree = { { , } and a positive integer , where is the node-set and is the edge set. Define partition ( ) = { , , … วินเทจ | ≤ , 1 ≤ ≤ n, where | | indicates the cardinal number of subset , i.e., the number of nodes in , and for each node ∈ , all its children nodes { | ∈children( )} must in a previous subset , where 1 ≤ » . Our goal is to find an optimal partition ( ) whose computational cost | ( )| is minimal. T V E k V E P V V1 V2 Vn Vi k i Vi Vi Vi v Vi c c v Vj j i P* V P* V Here subset consists of all nodes that will be computed at -th step (Fig. ), so | | ≤ indicates that we can compute nodes each step at most because the number of available threads is . The restriction “for each node ∈ , all its children nodes { | ∈children( )} must in a previous subset , where 1 ≤ < ” indicates that node can be processed only if all its child nodes are processed. Vi i 2e Vi k k k v Vi c c v Vj j i v DHS implementation We aim to find an optimal way to parallelize the computation of solving linear equations for each neuron model by solving the mathematical scheduling problem above. To get the optimal partition, DHS first analyzes the topology and calculates the depth ( ) สําหรับ nodes ทั้งหมด ∈ . Then, the following two steps will be executed iteratively until every node ∈ is assigned to a subset: (1) find all candidate nodes and put these nodes into candidate set . A node is a candidate only if all its child nodes have been processed or it does not have any child nodes. (2) if | | ≤ , i.e., the number of candidate nodes is smaller or equivalent to the number of available threads, remove all nodes in and put them into ในทางกลับกันลบ deepest nodes from and add them to subset . Label these nodes as processed nodes (Fig. ). After filling in subset , go to step (1) to fill in the next subset . d v v V v V Q Q k Q V*i k Q Vi 2d Vi วินาที + 1 Correctness proof for DHS After applying DHS to a neural tree = { , }, we get a partition ( ) = { , , … }, | | ≤ , 1 ≤ ≤ . Nodes in the same subset will be computed in parallel, taking steps to perform triangularization and back-substitution, respectively. We then demonstrate that the reordering of the computation in DHS will result in a result identical to the serial Hines method. T V E P V V1 V2 Vn Vi k i n วิน n The partition ( ) obtained from DHS decides the computation order of all nodes in a neural tree. Below we demonstrate that the computation order determined by ( ) satisfies the correctness conditions. ( ) is obtained from the given neural tree . Operations in DHS do not modify the tree topology and values of tree nodes (corresponding values in the linear equations), so the tree morphology and initial values of all nodes are not changed, which satisfies condition 1: the tree morphology and initial values of all nodes are identical to those in serial Hines method. In triangularization, nodes are processed from subset to . As shown in the implementation of DHS, all nodes in subset are selected from the candidate set , and a node can be put into only if all its child nodes have been processed. Thus the child nodes of all nodes in มีอยู่ใน { , , … }, meaning that a node is only computed after all its children have been processed, which satisfies condition 2: in triangularization, a node can be processed if and only if all its child nodes are already processed. In back-substitution, the computation order is the opposite of that in triangularization, i.e., from to . ตามที่แสดงไว้ก่อนหน้านี้ nodes เด็กของ nodes ทั้งหมดใน are in { , , … }, so parent nodes of nodes in are in { , , … }, which satisfies condition 3: in back-substitution, a node can be processed only if its parent node is already processed. P V P V P V T V1 สหภาพยุโรป Vi Q Q วิน V1 V2 Vi-1 Vn V1 Vi V1 V2 Vi-1 Vi Vi+1 Vi+2 Vn การพิสูจน์ประสิทธิภาพสําหรับ DHS The idea of the proof is that if there is another optimal solution, it can be transformed into our DHS solution without increasing the number of steps the algorithm requires, thus indicating that the DHS solution is optimal. สําหรับแต่ละส่วน ใน ( ), DHS moves (thread number) deepest nodes from the corresponding candidate set สอง . If the number of nodes in is smaller than , move all nodes from to เพื่อให้เป็นเรื่องง่ายเราแนะนํา , indicating the depth sum of deepest nodes in . All subsets in ( ) satisfy the max-depth criteria (Supplementary Fig. ): . We then prove that selecting the deepest nodes in each iteration makes an optimal partition. If there exists an optimal partition = { , , … } containing subsets that do not satisfy the max-depth criteria, we can modify the subsets in ( ) so that all subsets consist of the deepest nodes from and the number of subsets ( | ( )|) remain the same after modification. Vi P V k Qi Vi ชิ k Qi Vi Di k Qi P V 6a P(V) P*(V) V*1 V*2 V*s P* V Q P* V Without any loss of generalization, we start from the first subset not satisfying the criteria, i.e., . There are two possible cases that will make not satisfy the max-depth criteria: (1) | | < and there exist some valid nodes in that are not put to ; (2) | | = but nodes in are not the deepest nodes in . V*i V*i V*i k Qi V*i V * i k V*i k Qi For case (1), because some candidate nodes are not put to , these nodes must be in the subsequent subsets. As | | , we can move the corresponding nodes from the subsequent subsets to , which will not increase the number of subsets and make satisfy the criteria (Supplementary Fig. , top). For case (2), | | = , these deeper nodes that are not moved from the candidate set into must be added to subsequent subsets (Supplementary Fig. , bottom). These deeper nodes can be moved from subsequent subsets to through the following method. Assume that after filling , is picked and one of the -th deepest nodes is still in ดังนั้น will be put into a subsequent subset ( > ) เราย้ายครั้งแรก from to + , then modify subset + ออนไลน์ as follows: if | + | ≤ and none of the nodes in + is the parent of node , stop modifying the latter subsets. Otherwise, modify + as follows (Supplementary Fig. ): if the parent node of is in + , move this parent node to + ; else move the node with minimum depth from + to + . After adjusting , modify subsequent subsets + , + , … with the same strategy. Finally, move from to . V*i V*i < k V*i V*i 6b V*i k Qi V*i 6b V*i V*i v k v’ Qi v’ V*j j i v V*i V*i 1 V*i 1 V*i 1 k V*i 1 v V*i 1 6c v V*i 1 V*i 2 V*i 1 V*i 2 V * i V*i 1 V*i 2 V*j-1 v’ V*j V*i With the modification strategy described above, we can replace all shallower nodes in with the -th deepest node in and keep the number of subsets, i.e., | ( )| the same after modification. We can modify the nodes with the same strategy for all subsets in ( ) that do not contain the deepest nodes. Finally, all subsets ∈ ( ) can satisfy the max-depth criteria, and | ( )| does not change after modifying. V*i k Qi P * V P* V V*i P* V P* V In conclusion, DHS generates a partition ( ) และ Subsets ทั้งหมด ∈ ( ) satisfy the max-depth condition: . For any other optimal partition ( ) we can modify its subsets to make its structure the same as ( ), i.e., each subset consists of the deepest nodes in the candidate set, and keep | ( ) the same after modification. So, the partition ( ) obtained from DHS is one of the optimal partitions. P V วิน P V P* V P V P* V | P V GPU implementation and memory boosting To achieve high memory throughput, GPU utilizes the memory hierarchy of (1) global memory, (2) cache, (3) register, where global memory has large capacity but low throughput, while registers have low capacity but high throughput. We aim to boost memory throughput by leveraging the memory hierarchy of GPU. GPU employs SIMT (Single-Instruction, Multiple-Thread) architecture. Warps are the basic scheduling units on GPU (a warp is a group of 32 parallel threads). A warp executes the same instruction with different data for different threads . Correctly ordering the nodes is essential for this batching of computation in warps, to make sure DHS obtains identical results as the serial Hines method. When implementing DHS on GPU, we first group all cells into multiple warps based on their morphologies. Cells with similar morphologies are grouped in the same warp. We then apply DHS on all neurons, assigning the compartments of each neuron to multiple threads. Because neurons are grouped into warps, the threads for the same neuron are in the same warp. Therefore, the intrinsic synchronization in warps keeps the computation order consistent with the data dependency of the serial Hines method. Finally, threads in each warp are aligned and rearranged according to the number of compartments. 46 When a warp loads pre-aligned and successively-stored data from global memory, it can make full use of the cache, which leads to high memory throughput, while accessing scatter-stored data would reduce memory throughput. After compartments assignment and threads rearrangement, we permute data in global memory to make it consistent with computing orders so that warps can load successively-stored data when executing the program. Moreover, we put those necessary temporary variables into registers rather than global memory. Registers have the highest memory throughput, so the use of registers further accelerates DHS. Full-spine and few-spine biophysical models We used the published human pyramidal neuron . The membrane capacitance m = 0.44 μF cm-2, membrane resistance m = 48,300 Ω cm2, and axial resistivity a = 261.97 Ω cm. In this model, all dendrites were modeled as passive cables while somas were active. The leak reversal potential l = -83.1 mV. Ion channels such as Na+ and K+ were inserted on soma and initial axon, and their reversal potentials were Na = 67.6 mV, K = -102 mV respectively. All these specific parameters were set the same as in the model of Eyal, et al. , for more details please refer to the published model (ModelDB, access No. 238347). 51 c r r E E E 51 In the few-spine model, the membrane capacitance and maximum leak conductance of the dendritic cables 60 μm away from soma were multiplied by a spine factor to approximate dendritic spines. In this model, spine was set to 1.9. Only the spines that receive synaptic inputs were explicitly attached to dendrites. F F In the full-spine model, all spines were explicitly attached to dendrites. We calculated the spine density with the reconstructed neuron in Eyal, et al. . The spine density was set to 1.3 μm-1, and each cell contained 24994 spines on dendrites 60 μm away from the soma. 51 The morphologies and biophysical mechanisms of spines were the same in few-spine and full-spine models. The length of the spine neck neck = 1.35 μm and the diameter neck = 0.25 μm, whereas the length and diameter of the spine head were 0.944 μm, i.e., the spine head area was set to 2.8 μm2. Both spine neck and spine head were modeled as passive cables, with the reversal potential = -86 mV. The specific membrane capacitance, membrane resistance, and axial resistivity were the same as those for dendrites. L D El Synaptic inputs We investigated neuronal excitability for both distributed and clustered synaptic inputs. All activated synapses were attached to the terminal of the spine head. For distributed inputs, all activated synapses were randomly distributed on all dendrites. For clustered inputs, each cluster consisted of 20 activated synapses that were uniformly distributed on a single randomly-selected compartment. All synapses were activated simultaneously during the simulation. การจําลองการไหลเวียน synaptic ที่ขึ้นอยู่กับ AMPA และ NMDA เช่นเดียวกับในงานของ Eyal et al. การนําทาง AMPA ได้รับการจําลองเป็นฟังก์ชั่นการแสดงผลคู่และการนําทาง NMDA เป็นฟังก์ชั่นการแสดงผลคู่ที่ขึ้นอยู่กับแรงดันไฟฟ้า สําหรับรุ่น AMPA ฟังก์ชั่นที่เฉพาะเจาะจง rise and decay were set to 0.3 and 1.8 ms. For the NMDA model, rise and decay were set to 8.019 and 34.9884 ms, respectively. The maximum conductance of AMPA and NMDA were 0.73 nS and 1.31 nS. τ τ τ τ Background noise We attached background noise to each cell to simulate a more realistic environment. Noise patterns were implemented as Poisson spike trains with a constant rate of 1.0 Hz. Each pattern started at start = 10 ms and lasted until the end of the simulation. We generated 400 noise spike trains for each cell and attached them to randomly-selected synapses. The model and specific parameters of synaptic currents were the same as described in , except that the maximum conductance of NMDA was uniformly distributed from 1.57 to 3.275, resulting in a higher AMPA to NMDA ratio. t Synaptic Inputs Exploring neuronal excitability We investigated the spike probability when multiple synapses were activated simultaneously. For distributed inputs, we tested 14 cases, from 0 to 240 activated synapses. For clustered inputs, we tested 9 cases in total, activating from 0 to 12 clusters respectively. Each cluster consisted of 20 synapses. For each case in both distributed and clustered inputs, we calculated the spike probability with 50 random samples. Spike probability was defined as the ratio of the number of neurons fired to the total number of samples. All 1150 samples were simulated simultaneously on our DeepDendrite platform, reducing the simulation time from days to minutes. Performing AI tasks with the DeepDendrite platform Conventional detailed neuron simulators lack two functionalities important to modern AI tasks: (1) alternately performing simulations and weight updates without heavy reinitialization and (2) simultaneously processing multiple stimuli samples in a batch-like manner. Here we present the DeepDendrite platform, which supports both biophysical simulating and performing deep learning tasks with detailed dendritic models. DeepDendrite consists of three modules (Supplementary Fig. ): (1) an I/O module; (2) a DHS-based simulating module; (3) a learning module. When training a biophysically detailed model to perform learning tasks, users first define the learning rule, then feed all training samples to the detailed model for learning. In each step during training, the I/O module picks a specific stimulus and its corresponding teacher signal (if necessary) from all training samples and attaches the stimulus to the network model. Then, the DHS-based simulating module initializes the model and starts the simulation. After simulation, the learning module updates all synaptic weights according to the difference between model responses and teacher signals. After training, the learned model can achieve performance comparable to ANN. The testing phase is similar to training, except that all synaptic weights are fixed. 5 HPC-Net model Image classification is a typical task in the field of AI. In this task, a model should learn to recognize the content in a given image and output the corresponding label. Here we present the HPC-Net, a network consisting of detailed human pyramidal neuron models that can learn to perform image classification tasks by utilizing the DeepDendrite platform. HPC-Net has three layers, i.e., an input layer, a hidden layer, and an output layer. The neurons in the input layer receive spike trains converted from images as their input. Hidden layer neurons receive the output of input layer neurons and deliver responses to neurons in the output layer. The responses of the output layer neurons are taken as the final output of HPC-Net. Neurons between adjacent layers are fully connected. For each image stimulus, we first convert each normalized pixel to a homogeneous spike train. For pixel with coordinates ( ) in the image, the corresponding spike train has a constant interspike interval ISI( ) (in ms) which is determined by the pixel value ( ) as shown in Eq. ( ). x, y τ x, y p x, y 1 ในการทดลองของเราการจําลองสําหรับแต่ละกระตุ้นใช้เวลา 50 มม. รถไฟสปิกทั้งหมดเริ่มต้นที่ 9 + ISI ms and lasted until the end of the simulation. Then we attached all spike trains to the input layer neurons in a one-to-one manner. The synaptic current triggered by the spike arriving at time is given by τ t0 where is the post-synaptic voltage, the reversal potential syn = 1 mV, the maximum synaptic conductance max = 0.05 μS, and the time constant = 0.5 ms. v E g τ Neurons in the input layer were modeled with a passive single-compartment model. The specific parameters were set as follows: membrane capacitance m = 1.0 μF cm-2, membrane resistance m = 104 Ω cm2, axial resistivity a = 100 Ω cm, reversal potential of passive compartment l = 0 mV. c r r E The hidden layer contains a group of human pyramidal neuron models, receiving the somatic voltages of input layer neurons. The morphology was from Eyal, et al. , และ neurons ทั้งหมดถูกสร้างขึ้นโดยใช้สายเคเบิลพาสซีฟ ความจุของ membrane ที่เฉพาะเจาะจง m = 1.5 μF cm-2, membrane resistance m = 48,300 Ω cm2, axial resistivity a = 261.97 Ω cm, and the reversal potential of all passive cables l = 0 mV. Input neurons could make multiple connections to randomly-selected locations on the dendrites of hidden neurons. The synaptic current activated by the -th synapse of the -th input neuron on neuron ’s dendrite is defined as in Eq. ( ), where is the synaptic conductance, is the synaptic weight, is the ReLU-like somatic activation function, and is the somatic voltage of the -th input neuron at time . 51 c r r E k i j 4 gijk Wijk i t Neurons in the output layer were also modeled with a passive single-compartment model, and each hidden neuron only made one synaptic connection to each output neuron. All specific parameters were set the same as those of the input neurons. Synaptic currents activated by hidden neurons are also in the form of Eq. ( ). 4 การจัดอันดับภาพด้วย HPC-Net For each input image stimulus, we first normalized all pixel values to 0.0-1.0. Then we converted normalized pixels to spike trains and attached them to input neurons. Somatic voltages of the output neurons are used to compute the predicted probability of each class, as shown in equation , where is the probability of -th class predicted by the HPC-Net, is the average somatic voltage from 20 ms to 50 ms of the -เอาท์พุท neuron และ indicates the number of classes, which equals the number of output neurons. The class with the maximum predicted probability is the final classification result. In this paper, we built the HPC-Net with 784 input neurons, 64 hidden neurons, and 10 output neurons. 6 pi i i C Synaptic plasticity rules for HPC-Net Inspired by previous work , เราใช้กฎการเรียนรู้ตามเกรดเพื่อฝึก HPC-Net ของเราในการดําเนินการงานการจัดอันดับภาพ ฟังก์ชั่นการสูญเสียที่เราใช้ที่นี่คือ cross-entropy ให้ใน Eq. ( ), where is the predicted probability for class , indicates the actual class the stimulus image belongs to, = 1 if input image belongs to class , and = 0 if not. 36 7 pi i yi yi i yi When training HPC-Net, we compute the update for weight (the synaptic weight of the -th synapse connecting neuron to neuron ) at each time step. After the simulation of each image stimulus, is updated as shown in Eq. ( ): Wijk k i j Wijk 8 นี่คืออัตราการเรียนรู้เป็นค่าอัปเดตในเวลา , , are somatic voltages of neuron and respectively, is the -th synaptic current activated by neuron on neuron , its synaptic conductance, is the transfer resistance between the -th connected compartment of neuron on neuron ’s dendrite to neuron ’s soma, s = 30 ms, e = 50 ms เป็นเวลาเริ่มต้นและเวลาสิ้นสุดสําหรับการเรียนรู้สอดคล้องกัน สําหรับ neurons output ระยะเวลาความผิดพลาดสามารถคํานวณได้ตามที่แสดงใน Eq. ( ). For hidden neurons, the error term is calculated from the error terms in the output layer, given in Eq. ( ). t vj vi i j Iijk k i j gijk rijk k i j j t t 10 11 Since all output neurons are single-compartment, equals to the input resistance of the corresponding compartment, . Transfer and input resistances are computed by NEURON. การฝึกอบรมมินิแบทช์เป็นวิธีการทั่วไปในการเรียนรู้ลึกเพื่อให้ได้ความแม่นยําในการคาดการณ์ที่สูงขึ้นและเร่งการเข้ากันได้ DeepDendrite ยังสนับสนุนการฝึกอบรมมินิแบทช์ เมื่อฝึกอบรม HPC-Net ด้วยขนาดมินิแบทช์ batch, we make batch copies of HPC-Net. During training, each copy is fed with a different training sample from the batch. DeepDendrite first computes the weight update for each copy separately. After all copies in the current training batch are done, the average weight update is calculated and weights in all copies are updated by this same amount. N N ความแข็งแกร่งในการต่อต้านการโจมตีกับ HPC-Net To demonstrate the robustness of HPC-Net, we tested its prediction accuracy on adversarial samples and compared it with an analogous ANN (one with the same 784-64-10 structure and ReLU activation, for fair comparison in our HPC-Net each input neuron only made one synaptic connection to each hidden neuron). We first trained HPC-Net and ANN with the original training set (original clean images). Then we added adversarial noise to the test set and measured their prediction accuracy on the noisy test set. We used the Foolbox , เพื่อสร้างเสียงรบกวนต่อต้านด้วยวิธีการ FGSM ANN ได้รับการฝึกอบรมกับ PyTorch , and HPC-Net was trained with our DeepDendrite. For fairness, we generated adversarial noise on a significantly different network model, a 20-layer ResNet . The noise level ranged from 0.02 to 0.2. We experimented on two typical datasets, MNIST และแฟชั่น-MNIST . Results show that the prediction accuracy of HPC-Net is 19% and 16.72% higher than that of the analogous ANN, respectively. 98 99 93 100 101 95 96 Reporting summary ข้อมูลเพิ่มเติมเกี่ยวกับการออกแบบการวิจัยสามารถใช้ได้ใน linked to this article. Nature Portfolio Reporting Summary Data availability The data that support the findings of this study are available within the paper, Supplementary Information and Source Data files provided with this paper. The source code and data that used to reproduce the results in Figs. – are available at . The MNIST dataset is publicly available at . The Fashion-MNIST dataset is publicly available at . are provided with this paper. 3 6 https://github.com/pkuzyc/DeepDendrite http://yann.lecun.com/exdb/mnist https://github.com/zalandoresearch/fashion-mnist Source data Code availability The source code of DeepDendrite as well as the models and code used to reproduce Figs. – in this study are available at . 3 6 https://github.com/pkuzyc/DeepDendrite การอ้างอิง McCulloch, W. S. & Pitts, W. การคํานวณเชิงตรรกะของความคิดที่อยู่ภายใต้กิจกรรมประสาท. บูล. Math. Biophys. 5, 115–133 (1943). LeCun, Y., Bengio, Y. & Hinton, G. การเรียนรู้อย่างลึก ธรรมชาติ 521, 436–444 (2015) Poirazi, P., Brannon, T. & Mel, B. W. Arithmetic of subthreshold synaptic summation in a model CA1 pyramidal cell. , 977–987 (2003). Neuron 37 London, M. & Häusser, M. การคํานวณ Dendritic. Annu. Rev. Neurosci. 28, 503–532 (2005) Branco, T. & Häusser, M. The single dendritic branch as a fundamental functional unit in the nervous system. , 494–502 (2010). Curr. Opin. Neurobiol. 20 Stuart, G. J. & Spruston, N. การบูรณาการ Dendritic: 60 ปีของความก้าวหน้า. Nat. Neurosci. 18, 1713–1721 (2015). Poirazi, P. & Papoutsi, A. Illuminating dendritic function with computational models. , 303–321 (2020). Nat. Rev. Neurosci. 21 Yuste, R. & Denk, W. สกรู Dendritic เป็นหน่วยการทํางานพื้นฐานของการบูรณาการประสาท ธรรมชาติ 375, 682–684 (1995). Engert, F. & Bonhoeffer, T. Dendritic spine changes associated with hippocampal long-term synaptic plasticity. , 66–70 (1999). Nature 399 Yuste, R. Dendritic spines and distributed circuits. , 772–781 (2011). Neuron 71 Yuste, R. การแยกส่วนไฟฟ้าในลําไส้ dendritic. Annu. Rev. Neurosci. 36, 429–449 (2013). Rall, W. Branching dendritic trees and motoneuron membrane resistivity. , 491–527 (1959). Exp. Neurol. 1 Segev, I. & Rall, W. Computational study of an excitable dendritic spine. , 499–523 (1988). J. Neurophysiol. 60 Silver, D. et al. การครอบครองเกมของ Go กับเครือข่ายประสาทลึกและการค้นหาต้นไม้ Nature 529, 484–489 (2016). Silver, D. et al. A general reinforcement learning algorithm that masters chess, shogi, and go through self-play. , 1140–1144 (2018). Science 362 McCloskey, M. & Cohen, N. J. Catastrophic interference in connectionist networks: the sequential learning problem. , 109–165 (1989). Psychol. Learn. Motiv. 24 ฝรั่งเศส, R. M. การลืมอันตรายในเครือข่ายการเชื่อมต่อ แนวโน้ม Cogn. Sci. 3, 128–135 (1999) Naud, R. & Sprekeler, H. Sparse bursts optimize information transmission in a multiplexed neural code. , E6329–E6338 (2018). Proc. Natl Acad. Sci. USA 115 Sacramento, J., Costa, R. P., Bengio, Y. & Senn, W. microcircuits cortical dendritic approximate the backpropagation algorithm. in Advances in Neural Information Processing Systems 31 (NeurIPS 2018) (NeurIPS*,* 2018). Payeur, A., Guerguiev, J., Zenke, F., Richards, B. A. & Naud, R. Burst-dependent synaptic plasticity can coordinate learning in hierarchical circuits. , 1010–1019 (2021). Nat. Neurosci. 24 Bicknell, B. A. & Häusser, M. A synaptic learning rule for exploiting nonlinear dendritic computation. , 4001–4017 (2021). Neuron 109 Moldwin, T., Kalmenson, M. & Segev, I. The gradient clusteron: a model neuron that learns to solve classification tasks via dendritic nonlinearities, structural plasticity, and gradient descent. , e1009015 (2021). PLoS Comput. Biol. 17 Hodgkin, A. L. & Huxley, A. F. A quantitative description of membrane current and Its application to conduction and excitation in nerve. , 500–544 (1952). J. Physiol. 117 Rall, W. Theory of physiological properties of dendrites. , 1071–1092 (1962). Ann. N. Y. Acad. Sci. 96 Hines, M. L. & Carnevale, N. T. The NEURON simulation environment. , 1179–1209 (1997). Neural Comput. 9 Bower, J. M. & Beeman, D. in (eds Bower, J.M. & Beeman, D.) 17–27 (Springer New York, 1998). The Book of GENESIS: Exploring Realistic Neural Models with the GEneral NEural SImulation System Hines, M. L., Eichner, H. & Schürmann, F. Neuron splitting in compute-bound parallel network simulations enables runtime scaling with twice as many processors. , 203–210 (2008). J. Comput. Neurosci. 25 Hines, M. L., Markram, H. & Schürmann, F. การจําลองแบบคล้ายคลึงกันอย่างสมบูรณ์ของประสาทประสาทเดี่ยว J. Comput. Neurosci. 25, 439-448 (2008) Ben-Shalom, R., Liberman, G. & Korngreen, A. Accelerating compartmental modeling on a graphical processing unit. , 4 (2013). Front. Neuroinform. 7 Tsuyuki, T., Yamamoto, Y. & Yamazaki, T. Efficient numerical simulation of neuron models with spatial structure on graphics processing units. In (eds Hirose894Akiraet al.) 279–285 (Springer International Publishing, 2016). Proc. 2016 International Conference on Neural Information Processing Vooturi, D. T., Kothapalli, K. & Bhalla, U. S. Parallelizing Hines Matrix Solver in Neuron Simulations on GPU. In 388–397 (IEEE, 2017). Proc. IEEE 24th International Conference on High Performance Computing (HiPC) Huber, F. แก้ไขต้นไม้ที่มีประสิทธิภาพสําหรับ matrix hines บน GPU. พิมพ์ล่วงหน้าที่ https://arxiv.org/abs/1810.12742 (2018). Korte, B. & Vygen, J. ทฤษฎีการเพิ่มประสิทธิภาพการรวมและอัลกอริทึม 6 edn (Springer, 2018) Gebali, F. (Wiley, 2011). Algorithms and Parallel Computing Kumbhar, P. et al. CoreNEURON: An optimized compute engine for the NEURON simulator. , 63 (2019). Front. Neuroinform. 13 Urbanczik, R. & Senn, W. Learning by the dendritic prediction of somatic spiking. , 521–528 (2014). Neuron 81 Ben-Shalom, R., Aviv, A., Razon, B. & Korngreen, A. การเพิ่มประสิทธิภาพของรุ่นช่องไอออนโดยใช้อัลกอริทึมทางพันธุกรรมแบบขนานบนโปรเซสเซอร์กราฟิก J. Neurosci. วิธีการ 206, 183-194 (2012). Mascagni, M. A parallelizing algorithm for computing solutions to arbitrarily branched cable neuron models. , 105–114 (1991). J. Neurosci. Methods 36 McDougal, R. A. et al. 20 ปีของ modelDB และอื่น ๆ: การสร้างเครื่องมือการสร้างแบบจําลองที่จําเป็นสําหรับอนาคตของวิทยาศาสตร์ประสาท J. Comput. Neurosci. 42, 1-10 (2017). Migliore, M., Messineo, L. & Ferrante, M. Dendritic Ih บล็อกการรวมเวลาของอินพุต distal ที่ไม่ได้ซิงโครไนซ์ใน neurons pyramidal CA1 J. Comput. Neurosci. 16, 5–13 (2004) Hemond, P. et al. Distinct classes of pyramidal cells exhibit mutually exclusive firing patterns in hippocampal area CA3b. , 411–424 (2008). Hippocampus 18 Hay, E., Hill, S., Schürmann, F., Markram, H. & Segev, I. Models of neocortical layer 5b pyramidal cells capturing a wide range of dendritic and perisomatic active Properties. , e1002107 (2011). PLoS Comput. Biol. 7 Masoli, S., Solinas, S. & D’Angelo, E. Action potential processing in a detailed purkinje cell model reveals a critical role for axonal compartmentalization. , 47 (2015). Front. Cell. Neurosci. 9 Lindroos, R. et al. Basal ganglia neuromodulation over multiple temporal and structural scales—simulations of direct pathway MSNs investigate the fast onset of dopaminergic effects and predict the role of Kv4.2. , 3 (2018). Front. Neural Circuits 12 Migliore, M. et al. Synaptic clusters function as odor operators in the olfactory bulb. , 8499–8504 (2015). Proc. Natl Acad. Sci. USa 112 NVIDIA. . (2021). CUDA C++ Programming Guide https://docs.nvidia.com/cuda/cuda-c-programming-guide/index.html NVIDIA. CUDA C++ คู่มือการปฏิบัติที่ดีที่สุด. https://docs.nvidia.com/cuda/cuda-c-best-practices-guide/index.html (2021). Harnett, M. T., Makara, J. K., Spruston, N., Kath, W. L. & Magee, J. C. Synaptic amplification by dendritic spines enhances input cooperativity. , 599–602 (2012). Nature 491 Chiu, C. Q. et al. Compartmentalization of GABAergic inhibition by dendritic spines. , 759–762 (2013). Science 340 Tønnesen, J., Katona, G., Rózsa, B. & Nägerl, U. V. Spine neck plasticity regulates compartmentalization of synapses. , 678–685 (2014). Nat. Neurosci. 17 Eyal, G. et al. Human cortical pyramidal neurons: from spines to spikes via models. , 181 (2018). Front. Cell. Neurosci. 12 Koch, C. & Zador, A. The function of dendritic spines: devices subserving biochemical rather than electrical compartmentalization. , 413–422 (1993). J. Neurosci. 13 Koch, C. Dendritic spines. In (Oxford University Press, 1999). Biophysics of Computation Rapp, M., Yarom, Y. & Segev, I. ผลกระทบของกิจกรรมพื้นหลังเส้นใย paralel บนคุณสมบัติของสายเคเบิลของเซลล์ Purkinje cerebellar คอมพิวเตอร์ประสาท 4, 518-533 (1992). Hines, M. การคํานวณที่มีประสิทธิภาพของความสม่ําเสมอของประสาทสาขา Int. J. Bio-Med. Comput. 15, 69-76 (1984). Nayebi, A. & Ganguli, S. การป้องกันที่สร้างแรงบันดาลใจทางชีวภาพของเครือข่ายลึกจากโจมตีต่อต้านการต่อต้าน Preprint at https://arxiv.org/abs/1703.09202 (2017). Goddard, N. H. & Hood, G. การจําลองขนาดใหญ่โดยใช้ Genesis เปรียบเทียบ ในหนังสือของ Genesis: การสํารวจรูปแบบประสาทจริงด้วยระบบจําลองประสาททั่วไป (eds Bower James M. & Beeman David) 349-379 (Springer New York, 1998). Migliore, M., Cannia, C., Lytton, W. W., Markram, H. & Hines, M. L. Parallel network simulations with NEURON. , 119 (2006). J. Comput. Neurosci. 21 Lytton, W. W. et al. Simulation neurotechnologies for advancing brain research: parallelizing large networks in NEURON. , 2063–2090 (2016). Neural Comput. 28 Valero-Lara, P. et al. cuHinesBatch: Solving multiple Hines systems on GPUs human brain project. In 566–575 (IEEE, 2017). Proc. 2017 International Conference on Computational Science Akar, N. A. et al. Arbor—ห้องสมุดจําลองเครือข่ายประสาทที่มีรายละเอียดเชิงรูปร่างสําหรับสถาปัตยกรรมการคํานวณประสิทธิภาพสูงในปัจจุบัน ใน Proc. 27th Euromicro International Conference on Parallel, Distributed and Network-Based Processing (PDP) 274–282 (IEEE, 2019) Ben-Shalom, R. et al. NeuroGPU: Accelerating multi-compartment, biophysically detailed neuron simulations on GPUs. , 109400 (2022). J. Neurosci. Methods 366 Rempe, M. J. & Chopp, D. L. อัลกอริทึมการคาดการณ์การแก้ไขสําหรับการตอบสนองการแพร่กระจายสัดส่วนที่เกี่ยวข้องกับกิจกรรมประสาทในโครงสร้างสาขา SIAM J. Sci. Comput. 28, 2139-2161 (2006) Kozloski, J. & Wagner, J. An ultrascalable solution to large-scale neural tissue simulation. , 15 (2011). Front. Neuroinform. 5 Jayant, K. et al. Targeted intracellular voltage recordings from dendritic spines using quantum-dot-coated nanopipettes. , 335–342 (2017). Nat. Nanotechnol. 12 Palmer, L. M. & Stuart, G. J. Membrane potential changes in dendritic spines during action potentials and synaptic input. , 6897–6903 (2009). J. Neurosci. 29 Nishiyama, J. และ Yasuda, R. การคํานวณทางชีวเคมีสําหรับการพลาสติกโครงสร้างของกระดูกสันหลัง Neuron 87, 63–75 (2015). Yuste, R. & Bonhoeffer, T. Morphological changes in dendritic spines associated with long-term synaptic plasticity. , 1071–1089 (2001). Annu. Rev. Neurosci. 24 Holtmaat, A. & Svoboda, K. Experience-dependent structural synaptic plasticity in the mammalian brain. , 647–658 (2009). Nat. Rev. Neurosci. 10 Caroni, P., Donato, F. & Muller, D. Structural plasticity upon learning: regulation and functions. , 478–490 (2012). Nat. Rev. Neurosci. 13 Keck, T. et al. Massive restructuring of neuronal circuits during functional reorganization of adult visual cortex. , 1162 (2008). Nat. Neurosci. 11 Hofer, S. B., Mrsic-Flogel, T. D., Bonhoeffer, T. & Hübener, M. Experience leaves a lasting structural trace in cortical circuits. , 313–317 (2009). Nature 457 Trachtenberg, J. T. et al. Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. , 788–794 (2002). Nature 420 Marik, S. A., Yamahachi, H., McManus, J. N., Szabo, G. & Gilbert, C. D. Axonal dynamics of excitatory and inhibitory neurons in somatosensory cortex. , e1000395 (2010). PLoS Biol. 8 Xu, T. et al. Rapid formation and selective stabilization of synapses for enduring motor memories. , 915–919 (2009). Nature 462 Albarran, E., Raissi, A., Jáidar, O., Shatz, C. J. & Ding, J. B. การปรับปรุงการเรียนรู้มอเตอร์โดยการเพิ่มเสถียรภาพของสกรู dendritic ที่เกิดขึ้นใหม่ในเปลือกมอเตอร์ Neuron 109, 3298-3311 (2021). Branco, T. & Häusser, M. Gradients การบูรณาการ synaptic ใน dendrites ปิรามิดเซลล์ cortical เดียว Neuron 69, 885–892 (2011) Major, G., Larkum, M. E. & Schiller, J. Active properties of neocortical pyramidal neuron dendrites. , 1–24 (2013). Annu. Rev. Neurosci. 36 Gidon, A. et al. พลังงาน Dendritic และการคํานวณในชั้นมนุษย์ 2/3 เซลล์ประสาท cortical Science 367, 83–87 (2020) Doron, M., Chindemi, G., Muller, E., Markram, H. & Segev, I. Timed synaptic inhibition shapes NMDA spikes, influencing local dendritic processing and global I/O properties of cortical neurons. , 1550–1561 (2017). Cell Rep. 21 Du, K. et al. การยับยั้งชนิดเซลล์เฉพาะของศักยภาพเพลต dendritic ในประสาทประสาทการแสดงผลของกระดูกสันหลัง striatal. Proc. Natl Acad. Sci. USA 114, E7612-E7621 (2017). Smith, S. L., Smith, I. T., Branco, T. & Häusser, M. Dendritic spikes enhance stimulus selectivity in cortical neurons in vivo. , 115–120 (2013). Nature 503 Xu, N.-l et al. Nonlinear dendritic integration of sensory and motor input during an active sensing task. , 247–251 (2012). Nature 492 Takahashi, N., Oertner, T. G., Hegemann, P. & Larkum, M. E. Dendrites สารสกัดกร่อนที่ใช้งาน modulate การรับรู้. วิทยาศาสตร์ 354, 1587–1590 (2016). Sheffield, M. E. & Dombeck, D. A. การแพร่กระจายของแคลเซียมผ่านไปทั่วต้นไม้ dendritic การคาดการณ์คุณสมบัติของพื้นที่ ธรรมชาติ 517, 200–204 (2015). Markram, H. et al. การกู้คืนและการจําลองของไมโครวงจร neocortical เซลล์ 163, 456-492 (2015). Billeh, Y. N. et al. การบูรณาการระบบของข้อมูลโครงสร้างและฟังก์ชั่นในโมเดลหลายสแควร์ของกรงตาหลักของเมาส์ Neuron 106, 388–403 (2020) Hjorth, J. et al. microcircuits ของ striatum ในซิลิโคน. Proc. Natl Acad. Sci. USA 117, 202000671 (2020). Guerguiev, J., Lillicrap, T. P. & Richards, B. A. Towards deep learning with segregated dendrites. , e22901 (2017). elife 6 Iyer, A. et al. Avoiding catastrophe: active dendrites enable multi-task learning in dynamic environments. , 846219 (2022). Front. Neurorobot. 16 Jones, I. S. & Kording, K. P. Might a single neuron solve interesting machine learning problems through successive computations on its dendritic tree? , 1554–1571 (2021). Neural Comput. 33 Bird, A. D., Jedlicka, P. & Cuntz, H. Dendritic normalisation improves learning in sparsely connected artificial neural networks. , e1009202 (2021). PLoS Comput. Biol. 17 Goodfellow, I. J., Shlens, J. & Szegedy, C. Explaining and harnessing adversarial examples. In (ICLR, 2015). 3rd International Conference on Learning Representations (ICLR) Papernot, N., McDaniel, P. & Goodfellow, I. Transferability in machine learning: from phenomena to black-box attacks using adversarial samples. Preprint at (2016). https://arxiv.org/abs/1605.07277 Lecun, Y., Bottou, L., Bengio, Y. & Haffner, P. Gradient-based learning applied to document recognition. , 2278–2324 (1998). Proc. IEEE 86 Xiao, H., Rasul, K. & Vollgraf, R. Fashion-MNIST: a novel image dataset for benchmarking machine learning algorithms. Preprint at (2017). http://arxiv.org/abs/1708.07747 Bartunov, S. et al. Assessing the scalability of biologically-motivated deep learning algorithms and architectures. In (NeurIPS, 2018). Advances in Neural Information Processing Systems 31 (NeurIPS 2018) Rauber, J., Brendel, W. & Bethge, M. Foolbox: A Python toolbox to benchmark the robustness of machine learning models. In (2017). Reliable Machine Learning in the Wild Workshop, 34th International Conference on Machine Learning Rauber, J., Zimmermann, R., Bethge, M. & Brendel, W. Foolbox native: fast adversarial attacks to benchmark the robustness of machine learning models in PyTorch, TensorFlow, and JAX. , 2607 (2020). J. Open Source Softw. 5 Paszke, A. et al. PyTorch: An imperative style, high-performance deep learning library. In (NeurIPS, 2019). Advances in Neural Information Processing Systems 32 (NeurIPS 2019) He, K., Zhang, X., Ren, S. & Sun, J. การเรียนรู้ที่เหลือลึกสําหรับการรับรู้ภาพ ใน Proc. 2016 IEEE Conference on Computer Vision and Pattern Recognition (CVPR) 770-778 (IEEE, 2016) การยอมรับ The authors sincerely thank Dr. Rita Zhang, Daochen Shi and members at NVIDIA for the valuable technical support of GPU computing. This work was supported by the National Key R&D Program of China (No. 2020AAA0130400) to K.D. and T.H., National Natural Science Foundation of China (No. 61088102) to T.H., National Key R&D Program of China (No. 2022ZD01163005) to L.M., Key Area R&D Program of Guangdong Province (No. 2018B030338001) to T.H., National Natural Science Foundation of China (No. 61825101) to Y.T., Swedish Research Council (VR-M-2020-01652), Swedish e-Science Research Centre (SeRC), EU/Horizon 2020 No. 945539 (HBP SGA3), and KTH Digital Futures to J.H.K., J.H., and A.K., Swedish Research Council (VR-M-2021-01995) and EU/Horizon 2020 no. 945539 (HBP SGA3) to S.G. and A.K. Part of the simulations were enabled by resources provided by the Swedish National Infrastructure for Computing (SNIC) at PDC KTH partially funded by the Swedish Research Council through grant agreement no. 2018-05973. กระดาษนี้สามารถใช้ได้ภายใต้ใบอนุญาต CC by 4.0 Deed (Attribution 4.0 International) กระดาษนี้สามารถใช้ได้ภายใต้ใบอนุญาต CC by 4.0 Deed (Attribution 4.0 International)