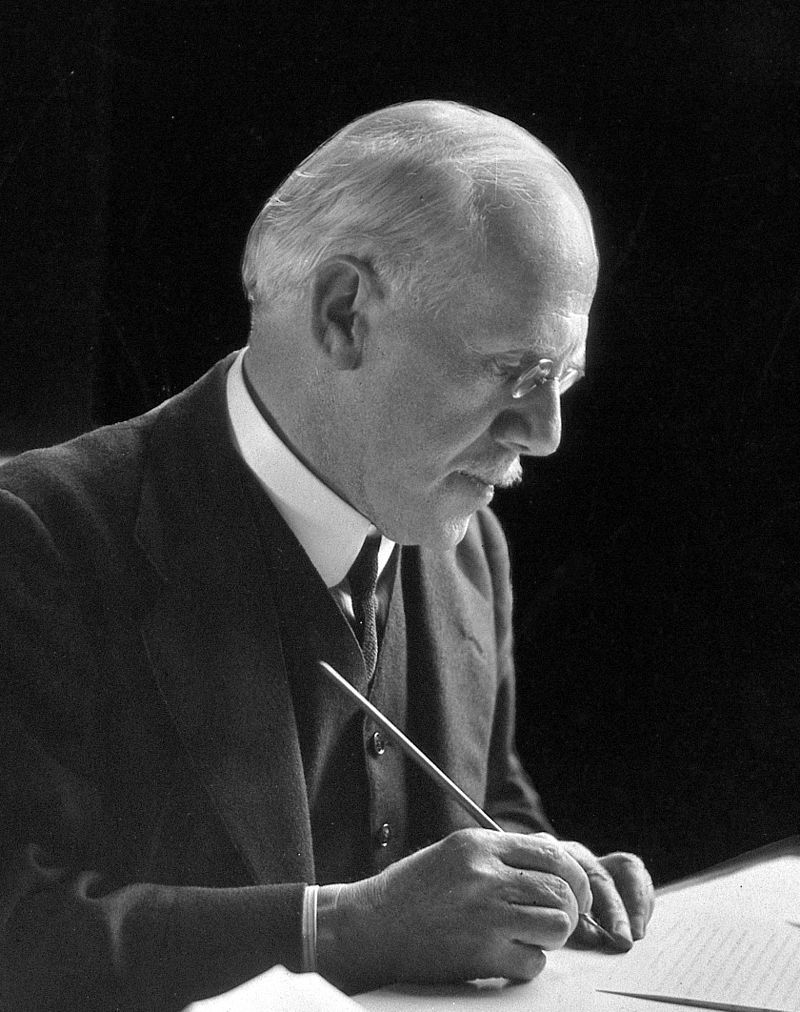Bacteria, by George Newman is part of the HackerNoon Books series. You can jump to any chapter in this book here . Chapter VIII: BACTERIA AND DISEASE CHAPTER VIII. BACTERIA AND DISEASE Probably the most universally known fact respecting bacteria is that they are related in some way to the production of disease. Yet we have seen that it was not as disease-producing agents that they were first studied. Indeed, it is only within comparatively the latest period of the two centuries during which they have been more or less under observation that our knowledge of them as causes of disease has assumed any exactitude or general recognition. Nor is this surprising, for although an intimate relationship between fermentation and disease had been hinted at in the middle of the seventeenth century, it was not till the time of Pasteur that the bacterial cause of fermentation was experimentally and finally established. In the middle of the seventeenth century men learned, through the eyes of Leeuwenhoek, that drops of water contained "moving animalcules." A hundred years later Spallanzani demonstrated the fact that putrefaction and fermentation were set up in boiled vegetable infusions when outside air was admitted, but when it was withheld from these boiled infusions no such change occurred. Almost a hundred years more passed before the epoch-making work of Tyndall and Pasteur, who separated these putrefactive germs from the air. Quickly following in their footsteps came Davaine and Pollender, who found in the blood of animals suffering from anthrax the now well-known specific and causal bacillus of that disease. Improvements in the microscope and in methods of cultivation (Koch's plate method in particular) soon brought an army of zealous investigators into the field, and during the last twenty years first this disease and then that have been traced to a bacterial origin. We may summarise the vast mass of historical, physiological, and pathological research extending from 1650 to 1898 in three great periods: the period of detection of living, moving cells (Leeuwenhoek and others in the seventeenth century); the period of the discovery of their close relationship to fermentation and putrefaction (Spallanzani, Schulze, Schwann, in the eighteenth century); and, thirdly, the period of appreciation of the rôle of bacteria in the economy of nature and in the production of disease (Tyndall, Pasteur, Lister, Koch, in the nineteenth). But we must look less cursorily at the growth of the idea of bacteria causing disease. More than two hundred years ago Robert Boyle (1627–91), the philosopher, who did so much towards the foundation of the present Royal Society, wrote a learned treatise on The Pathological Part of Physic. He was one of the earliest scientists to declare that a relationship existed between fermentation and disease. When more accurate knowledge was attained respecting fermentation, great advance was consequently made in the etiology of disease. The preliminary discoveries of Fuchs and others between 1840 and 1850 had relation to the existence in diseased tissues of a large number of bacteria. But this was no proof that such germs such diseases. It was not till Davaine had inoculated healthy animals with bacilli from the blood of an anthrax carcass, and had thus produced the disease, that reliance could be placed upon that bacillus as the of anthrax. Too much emphasis cannot be laid upon this idea, that unless a certain organism in healthy tissues the disease in question, it cannot be considered as proven that the particular organism is related to the disease as cause to effect. In order to secure a standard by which all investigators should test their results, Koch introduced four postulates. Until each of the four has been fulfilled, the final conclusion respecting the causal agent must be considered . The postulates are as follows: caused vera causa produces sub judice ( ) The organism must be demonstrated in the circulation or tissues of the diseased animal. a ( ) The organism thus demonstrated must be cultivated in artificial media outside the body, and successive generations of a of that organism must be obtained. b pure culture ( ) Such pure cultures must, when introduced into a healthy and susceptible animal, produce the specific disease. c ( ) The organism must be found and isolated from the circulation or tissues of the inoculated animal. d It is evident that there are some diseases—for example, cholera, leprosy, and typhoid—which are not communicable to lower animals, and therefore their virus cannot be made to fulfil postulate ( ). In such cases there is no choice. They cannot be classified along with tubercle and anthrax. Bacteriologists have little doubt that Hansen's bacillus of leprosy is the cause of that disease, yet it has not fulfilled postulates ( ) and ( ). Nor has the generally accepted bacillus of typhoid fulfilled postulate ( ), yet by the majority it is accepted as the agent in producing typhoid. Hence it will be seen that, though there is an academical classification of causal pathogenic bacteria according as they respond to Koch's postulates, yet nevertheless, there are a number of pathogenic bacteria which are looked upon as causes of disease provisionally. c b c c provisionally Anthrax and tubercle, with perhaps the organisms of suppuration, tetanus, plague, and actinomycosis, stand in the first order of pathogenic germs. Then comes a group awaiting further confirmation. It includes the organisms related to typhoid, cholera, malaria, leprosy, diarrhœa,and pneumonia. Then comes in a third category, a long list of diseases, such as scarlet fever, small-pox, rabies, and others too numerous to mention, in which the nature of the causal agent is still unknown. Hence it must not be supposed that every disease has its germ, and without a germ there is no disease. Such universal assertions, though not uncommonly heard, are devoid of accuracy. In the production of bacterial disease there are two factors. First, there is the body tissue of the individual; secondly, there is the specific organism. Whatever may be said hereinafter with regard to the power of micro-organisms to cause disease, we must understand one cardinal point, namely, that bacteria are never more than causes, for the nature of disease depends upon the behaviour of the organs or tissues with which the bacteria or their products meet (Virchow). Fortunately for a clear conception of what "organs and tissues" mean, these have been reduced to a common denominator, the cell. Every living organism, of whatever size or kind, and every organ and tissue in that living organism, contains and consists of cells. Further, these cells are composed of organic chemical substances which are not themselves alive, but the mechanical arrangement of which determines the direction and power of their organic activity and of their resistance to the specific agents of disease. With these facts clearly before us, we may hope to gain some insight into the reasons for departure from health. The normal living tissues have an inimical effect upon bacteria. Saprophytic bacteria of various kinds are normally present on exposed surfaces of skin or mucous membrane. Tissues also which are dead or depressed in vitality from injury or previous disease, but which are still in contact with the tissues, afford an excellent nidus for the growth of bacteria. Still these have not the power, unless specific, to thrive in the normal living tissue. It has been definitely shown that the blood fluids of the body have in their fresh state the germicidal power ( ) which prevents bacteria from flourishing in them. Such action does undoubtedly depend in measure upon the number of germs as well as their quality, for the killing power of blood and lymph must be limited. alexines Buchner has pointed out that the antagonistic action of these fluids depends in part possibly upon phagocytosis, but largely upon a chemical condition of the serum. The blood, then, is no friend to intruding bacteria. Its efforts are to a certain extent seconded by the lymphoid tissue throughout the body. Rings of lymphoid tissue surround the oral openings of the trachea (windpipe) and œsophagus (gullet); the tonsils are masses of lymphoid tissue. Composed as it is of cells having a germicidal influence when in health, the lymphoid tissue may afford formidable obstruction to intruding germs. All the foregoing points in one direction, namely, that if the tissues are maintained in sound health, they form a very resistant barrier against bacteria. But we know from experience that a full measure of health is not often the happy condition of human tissues; we have, in short, a variety of circumstances which, as we say, the individual to disease. One of the commonest forms of predisposition is that due to . Probably it is true that what are known as hereditary diseases are due far more to a hereditary predisposition than to any transmission of the virus itself in any form. predispose heredity predisposes the tissues to form a nidus for bacteria; frequently act in the same way. Damp soils must be held responsible for many disasters to health, not directly, but indirectly, by predisposition; dusty trades and injurious occupations have a similar effect. Any one of these three different influences may in a variety of ways affect the tissues and increase their susceptibility to disease. Not infrequently we may get them combined. For example, the following is not an unlikely series of events terminating in consumption (tuberculosis of the lungs):—( ) The individual is predisposed by inheritance to tuberculosis; ( ) an ordinary chronic catarrh, which lowers the resisting power of the lungs, may be contracted; ( ) the epithelial collections in the air vesicles of the lung— , dead matter attached to the body—afford an excellent nidus for bacteria; ( ) owing to occupation, or personal habits, or surroundings, the patient comes within a range of tubercular infection, and the specific bacilli of tubercle gain access to the lungs. The result, it is needless to state, will be a case of consumption more or less acute according to environment and treatment. Antecedent disease conditions of environment or personal habits a b c i. e. d The by which organisms gain the vantage-ground afforded by the depressed tissues are various, and next to the maintenance of resistant tissues they call for most attention from the physician and surgeon. It is in this field of preventive medicine—that is to say, preventing infective matter from ever entering the tissues at all—that science has triumphed in recent years. It is, in short, applied bacteriology, and therefore claims consideration in this place. channels of infection 1. By this term may be understood the actual transmission from the mother to the unborn child of the specific virus of the disease. That such a conveyance may occur is generally admitted by pathologists, but it is impossible to enter fully into the matter in such a book as the present. Summarily we may say that, though this sort of transmission is possible, it is not frequent, nor is disease appreciably spread through such a channel. Sixty per cent. of consumptives, it has been estimated, have tuberculous progenitors, and this is the highest figure. Many would be justified from experience in placing it at half that number. Pure Heredity. 2. , or inserting virus through a broken surface of skin, is itself a sufficiently obvious mode of infection to call for little comment. Yet it is under this heading that a word must be said of that remarkable application of preventive medicine known as the antiseptic treatment of wounds. When Lord Lister was Professor of Surgery in Glasgow, he was impressed with the greatness of the evil of putrefaction in wounds, which was caused, not by the oxygen of the air, as Liebig had declared, but by the entrance into the wound of fermentative organisms from the air. This was demonstrated by Pasteur, who pointed out that they could not arise in the wound. Hence it appeared to Lister that these fermentative bacteria which produce putrefaction in wounds must either be kept out of the wound altogether, or killed, or their action prevented, in the wound. To keep air away from wounds is an almost impossible task, and thus it came about that wounds were dressed with a solution of carbolic acid. Inoculation de novo From time to time examples occur of bacterial disease being directly inoculated in wounds made with polluted instruments, or in cuts made by contaminated broken glass, or in gunshot wounds. Tetanus is, of course, one of the most marked examples. 3. is a term which has suffered from the many ways in which it has been used. Defined shortly and most simply, we should say a disease is when it can be "caught" by contact, through the unbroken surfaces, between diseased and healthy persons. Ringworm is an example, and there are many others. Contagion contagious 4. The recent Royal Commission on Tuberculosis has collected a large mass of evidence in support of the view that tubercle may be spread by articles of food. Milk and meat from tuberculous animals naturally come in for the largest amount of condemnation. To these matters we refer elsewhere. The Alimentary Canal: Food. 5. The air may become infected with germs of disease from dusty trades, dried sputum, etc. If such infected air be inhaled, pathogenic results will follow, especially if the bacteria are present in sufficient numbers, or meet with devitalised, and therefore non-resisting, tissues. The Respiratory Tract: Air. These, then, are the five possible ways in which germs gain access to the body tissues. The question now arises, How do bacteria, having obtained entrance, set up the process of disease? For a long time pathologists looked upon the action of these microscopic parasites in the body as similar to, if not identical with, the larger parasites sometimes infesting the human body. Their work was viewed as a devouring of the tissues of the body. Now, it is well known that, however much or little of this may be done, the specific action of pathogenic bacteria is of a different nature. It is twofold. We have the action of the bacteria themselves, and also of their products or toxins. In particular diseases, now one and now the other property comes to the front. In bacterial diseases affecting or being transmitted mostly by the blood, it is the toxins which act chiefly. The convenient term is applied to those conditions in which there has been a multiplication of living organisms after they have entered the body, the word indicating a condition of poisoning brought about by their products. It will be apparent at once that we may have both these conditions present, the former before the latter, and the latter following as a direct effect of the former. infection intoxication Until intoxication occurs there may be few or no symptoms, but directly enough bacteria are present to produce in the body certain poisons in sufficient amount to result in more or less marked tissue change, then the symptoms of that tissue change appear. This period of latency between infection and the appearance of the disease is known as the . Take typhoid, for example. A man drinks a typhoid-polluted water. For about fourteen days the bacilli are making headway in his body without his being aware of it. But at the end of that incubation period the signs of the disease assert themselves. Professor Watson Cheyne and others have maintained that there is some exact proportion between the number of bacteria gaining entrance and the length of the incubation period. incubation period Speaking generally, we may note that pathogenic bacteria divide themselves into two groups: those which, on entering the body, pass at once, by the lymph or blood stream, to all parts of the body, and become more and more diffused throughout the blood and tissues, although in some cases they settle down in some spot remote from the point of entrance, and produce their chief lesions there. Tubercle and anthrax would be types of this group. On the other hand, there is a second group, which remain almost absolutely local, producing only little reaction around them, never passing through the body generally, and yet influencing the whole body eventually by means of their ferments or toxins. Of such the best representatives are tetanus and diphtheria. The local site of the bacteria is, in this case, the local manufactory of the disease. Whilst the mere bodily presence of bacteria may have mechanical influence injurious to the tissues (as in the small peripheral capillaries in anthrax), or may in some way act as a foreign body and be a focus of inflammation (as in tubercle), the real disease-producing action of pathogenic bacteria depends upon the (toxins) formed directly or indirectly by them. Though within recent years a great deal of knowledge has been acquired about the formation of these bodies, their exact nature is not known. They are allied to albuminous bodies and proteoses, and are frequently described as tox-albumens. It may be found, after all, that they are not of a proteid nature. chemical poisons Sidney Martin has pointed out that there is much that is analogous between the production of toxins and the production of the bodies of digestion. Just as ferments are necessary in the intestine to bring about a change in the food by which the non-soluble albumens shall be made into soluble peptones and thus become absorbed through the intestinal wall, so also a ferment may be necessary to the production of toxins. Such ferments have not as yet been isolated, but their existence in diphtheria and tetanus is, as we have seen, extremely likely. However that may be, it is now more or less established that there are two kinds of toxic bodies, differing from each other in their resistance to heat. It may be that the one most easily destroyed by heat is a ferment and possibly an originator of the other. A second division which has been suggested for toxic bodies, and to which reference has been made, is and , according to whether or not the poison exists within or without the body of the bacillus. intracellular extracellular Lastly, we may turn to consider the action of the toxins on the individual in whose body-fluids they are formed. It is hardly necessary to say that any action which bacteria or toxins may have will depend upon their virulence, in some measure upon their number, and not a little upon the channel of infection by which they have gained entrance. It could not be otherwise. If the virulence is attenuated, or if the invasion is very limited in numbers, it stands to reason that the pathogenic effects will be correspondingly small or absent. The influence of the toxins is twofold. In the first place (i.) they at the site of their formation, or at distant points by absorption. There is inflammation with marked cell-proliferation, and this is, more or less rapidly, followed by a specific cell-poisoning. The former change may be accompanied by exudation, and simulate the early stages of abscess formation; the latter is the specific effect, and results, as in leprosy and tubercle, in infective nodules. The site in some diseases, like typhoid (intestinal ulceration, splenic and mesenteric change) or diphtheria (membrane in the throat), may be definite and always the same. But, on the other hand, the site may depend upon the point of entrance, as in tetanus. act locally upon the tissues The distant effects of the toxin are due to absorption, but what controls its action it is impossible to say. We only know that we do find pathological conditions in certain organs at a distance and without the presence of bacteria. We have a parallel in the action of drugs; for example, a drug may be given by the mouth and yet produce a rash in some distant part of the body. In the second place (ii.) toxins produce . Fever and many of the from bacterial action must thus be classified. We have, it is true, the chemical symptoms of the pathological tissue change, for example, the large spleen of anthrax or the obstruction from diphtheritic membrane. But, in addition to these, we have general symptoms, as fever, in which after death no tissue change can be formed. toxic symptoms nervous conditions resulting We may now consider briefly some of the more important types of disease produced by bacteria: 1. Tuberculosis. As far back as 1794 Baillie drew attention to the grey miliary nodules occurring in tuberculous tissue which gave rise to the term "tubercles." This preliminary matter was confirmed by Bayle in 1810. In 1834 Laennec described all caseous deposits as "tubercles," insisting upon four varieties: (1) Miliary, which were about the size of millet seeds, and in groups; (2) Crude, miliary tubercles in yellow masses; (3) Granular, similar to the last, but scattered; (4) Encysted, a hard mass of crude tubercle with a fibrous or semi-cartilaginous capsule. The tubercle possesses in many cases a special structure, and certain cell-forms frequently occur in it and give it a characteristic appearance. The central part of the tubercle usually contains with numerous nuclei. The uninuclear cells are partly lymphoid, partly large epithelial or endothelial cells; these are called cells. giant cells epithelioid It was not till 1865 that the specific nature of tuberculosis was asserted by Villemin. Burdon Sanderson (1868–69) in England confirmed his work, and it was extended by Connheim, who a few years later laid down the principle that all is tubercular which by transference to properly constituted animals is capable of inducing tuberculosis, and nothing is tubercular unless it has this capability. Klebs (1877) and Max Schiller (1880) described masses of living cells or micrococci in many tuberculous nodules in the diseased synovial membrane and in lupus skin. In 1881 Toussaint declared he had cultivated from the blood of tubercular animals and from tubercles an organism which was evidently a micrococcus, and in the same year Aufrecht stated that the centre of a tubercle contained small micrococci, diplococci, and some rods. But it was not till the following year, 1882, that Koch discovered and demonstrated beyond question the specific . Bacillus tuberculosis It is now held to be absolutely proved that the introduction of the bacillus, or its spores or products, is the one and only essential agent in the production of tuberculosis. Its recognised manifestations are as follows: The disease may occur generally throughout the body or locally in the suprarenal capsules, prostate, intestine, larynx, membranes of the heart, bones, ovaries, pleura, kidneys, spleen, testicles, Fallopian tubes, uterus, etc. We may summarise the history of the pathology of tubercle thus: 1794. Baillie drew attention to grey miliary nodules occurring in tuberculosis, and called them "tubercles." 1834. Laennec described four varieties: ; ; ; . miliary crude granular encysted 1843. Klencke produced tuberculosis by intravenous injection of tubercular giant cells. 1865. Villemin demonstrated infectivity of tubercular matter by inoculation of discharges; Connheim, Armanni, Burdon Sanderson, Wilson Fox, and others showed that nothing but tubercular matter could produce tuberculosis. 1877. Living cells were found in tubercles, "micrococci" (Klebs, Toussaint, Schiller). 1882. Koch isolated and described the specific bacillus, and obtained pure cultivations (1884). , 1882. Delicate cylindrical rods, measuring 1.5–4 micromillimetres in length and about .2 µ in breadth; non-motile. Many are straight with rounded ends; others are slightly curved. They are usually solitary, but may occur in pairs, lying side by side or in small masses. They are chiefly found in fresh tubercles, more sparingly in older ones. Some lie within the giant cells; others lie outside; shorter in tissue sections of bovine tuberculosis, but longer in the milk (Crookshank). The Bacillus of Koch When stained they appear to be composed of irregular cubical or spherical granules within a faintly stained sheath. In recent lesions the protoplasm appears more homogeneous, and takes on the segmented or beaded character only in old lesions, pus, or sputum. Morphological differences are found under different circumstances, and within limits variation occurs according to the environment. Koch inoculated with tubercular matter from an infected lymphatic gland of a guinea-pig, and noticed the first signs of growth in ten or twelve days in the form of whitish, scaly patches. These enlarged and coalesced with neighbouring patches, forming white, roughened, irregular masses. Nocard and Roux showed that by adding 5/8 per cent. of glycerine to the media commonly used in the laboratory, such as nutrient agar or broth, the best growth is obtained. Cultivation on Various Media. solid blood serum On or abundant growth appears at the end of seven or eight days. By continuous sub-culture on glycerine agar the virulence of the bacillus is diminished. But in fifteen days after inoculation of the medium the culture equals in extent a culture of several weeks' age on blood serum. glycerine broth glycerine agar Sub-cultures from glycerined media will grow in ordinary broth without glycerine (Nocard, Roux, Crookshank). In to which a piece of boiled white of egg was added Klein obtained copious growths, and found that continued sub-culturing upon this medium also lessens the virulence. alkaline broth —On glycerine agar minute white colonies appear in about six days, raised and isolated, and coalescing as time advances, forming a white lichenous growth, fully developed in about two months. Description of Cultivations: On glycerine broth a copious film appears on the surface of the liquid, which if disturbed falls to the bottom of the flask as a deposit. In very old cultivations spore-like bodies can be observed both in stained and unstained preparations, but neither the irregular granules within the capsule nor the unstained spaces between the granules are spores (Babes and Crookshank). That the bacilli possess spores is believed on account of the following facts: Spore Formation. 1. That tubercular sputum, when thoroughly dried, maintains its virulent character (Koch, Schill, Fischer, etc.). No sporeless bacillus is known which can survive through drying. 2. That tubercular matter and cultures survive temperature up to 100° C. Non-spore-bearing bacilli and micrococci are killed by being exposed for five minutes to a temperature of 65–70° C., whereas spores of other bacilli withstand much higher temperatures. 3. Tubercular sputum distributed in salt solution does not lose its virulence by being kept at 100° C. for one or two minutes; sporeless bacilli certainly would (Klein). 4. A solution of per-chloride of mercury does not kill the tubercle bacilli, as it does sporeless bacilli (Lingard and Klein). Koch and many bacteriologists have declared the bacillus to be a "true parasite." Koch based this view upon the belief which he entertained that the bacillus can grow only between 30° C. and 41° C., and therefore in temperate zones is limited to the animal body and can originate only in an animal organism. "They are," he said, "true parasites, which cannot live without their hosts. They pass through the whole cycle of their existence in the body." But at length Koch and others overcame the difficulties and grew the bacillus as a saprophyte. Schottelius has observed that tubercle bacilli taken from the lung of phthisical persons buried for years still retains its virulence and capability of producing tuberculosis upon inoculation. He further shows that tubercular lung kept in soil (enclosed in a box) shows a marked rise in temperature. Klein quotes these experiments as indications that "tubercle bacilli are not true parasites, but belong to the ecto genic microbes which can live and thrive independent of a living host." It has now been abundantly proved that the bacillus of tuberculosis is capable of accommodating itself to circumstances much less favourable than had been supposed, especially as regards temperature. 30–41° C. have been laid down by Koch as the limits of temperature at which the bacillus will grow in culture medium outside the body. The generally accepted temperatures as most favourable to the growth of the bacillus are between 36° C. and 38° C. Temperature of Growth of Bacillus. Sir Hugh Beevor, however, was able to grow the bacillus upon glycerine agar at 28° C. (82° F.), obtaining an ample culture which developed somewhat more slowly than on blood serum, and to a less extent than at 37° C. In both Beevor succeeded in growing the bacillus at a lower temperature even than on agar, viz., at a temperature rarely above 60° F. Sheridan Delépine and others have also been successful in obtaining growths at room temperature both in summer and winter. Although, speaking generally, there is an actual cessation of growth at low temperature, the bacillus may be exposed to very low temperatures for a considerable time without losing its power of again becoming active when returned to a favourable environment (Woodhead). All four of Koch's postulates have been fulfilled in the case of . Hence we are dealing with the specific cause of the disease. Yet, whilst this is so, we may usefully ask ourselves: does the bacillus set up the changes in normal tissues which result in tubercular nodules? In arriving at a solution of this problem we are materially aided if we bear in mind the fact that such an organism in healthy tissues has a double effect. The Relation of the Bacillus to the Disease. Bacillus tuberculosis How First, there is an ordinary inflammatory irritation, and secondly, there is a specific change set up by the toxins of the bacillus. Directly the invading bacilli find themselves in a favourable nidus they commence multiplication. In three or four days this acts as an irritant upon the surrounding connective-tissue cells, which proliferate, and become changed into large cells known as . At the periphery of this collection of epithelioid cells we have a congested area. This change has been accomplished by the presence of the bacilli themselves. epithelioid cells The production of their specific poisons changes the epithelioid cells in the centre of the nodule, some of which become fused together, whilst others expand and undergo division of nucleus. By this means we obtain a series of large multi-nucleated cells named . If the disease is very active, these soon caseate and break down in the centre. In a limb we get a discharge; in a lung we get an expectoration. Both discharge and expectoration arise from a breaking down of the new cell formation. giant cells Previously to breaking down we have in a fully developed nodule healthy tissue, inflammatory zone, epithelioid cells, giant cells, containing nuclei and bacilli. The sputum or the discharge will, during the acute stage of the disease at all events, contain countless numbers of the bacilli, which may thus be readily detected, and their presence used as evidence of the disease. It is obvious that if the centre of the nodule degenerates and comes away as discharge a cavity will be left behind. By degrees this small cavity may become a very large one, as is frequently the case in the lung, which particularly lends itself to such a condition. Hence, though at the outset a tubercular lung is solid, at the end it is hollow. The exact period of giant-cell formation depends on the rapidity of the formative processes. Thus different conditions occur. Inside the giant cells the bacilli are arranged in relation to the nuclei in one of three ways: ( ) polar, ( ) zonal, or ( ) mixed. The breaking down of the nodule is partly due to the cell-poisons, and partly because the nodule is non-vascular, owing to the fact that new capillaries cannot grow into the dense nodule, and the old ones are all occluded by the growth of the nodule. a b c From the local foci of disease the tubercle process spreads chiefly by three channels: ( ) , affecting particularly the glands. Thus we get tuberculosis set up in the bronchial, tracheal, mediastinal, and mesenteric glands, and it is so frequently present as to be a characteristic of the disease. This is the common method of dissemination in the body. a By the lymphatics ( ) , by means of which bacilli may be carried to distant organs. b By the blood-vessels ( ) , infective giant-cell systems encroaching upon neighbouring tissues, or discharge from lungs or bronchial glands obtaining entrance to the gullet and thus setting up intestinal disease also. c By continuity of tissues It has been abundantly proved that the respiratory and digestive systems are principally affected by Koch's bacillus. Wherever the bacilli are arrested, they excite formation of granulations or miliary tubercular nodules, which increase and eventually coalesce. The lymphatic glands which collect the lymph from the affected region are the earliest affected, always the nearest first, and then the disease appears to be appreciably stopped on its invading march. Each lymphatic gland acts as a temporary barrier to progress until the disease has broken its structure down. It remains local, in spite of increase in number and importance of the foci of disease, as long as the bacilli have not gained access to the blood stream. Koch, Crookshank, and Herroun, Hunter, and others have isolated products from pure cultures of the tubercle bacillus. These have comprised chiefly , , and various . Koch's observations led him to suppose that in pure cultures of tubercle a substance appeared having healing action on tuberculosis, and an extract of this in glycerine he termed "tuberculin." It was made as follows: A veal broth containing peptone and glycerine was inoculated with a pure culture of the bacillus and incubated at 38° C. for six or eight weeks. An abundant growth with copious film formation appeared. The culture was then concentrated by evaporation over a water-bath until reduced to about one-tenth of its volume. Toxins and Tuberculin. albumoses alkaloids extractives Flask used in the Preparation of Tuberculin The announcements in 1890 and 1891 to the effect that a "cure" had been discovered for consumption will be remembered. The hopes thus raised were unfortunately not to be realised. Koch advocated injections of this tuberculin in cases of skin tubercle (lupus) and consumptive cases. In many of these benefit was apparently derived, but its general application was not founded upon any substantial basis. Dead tissue, full of bacilli, could not thus be got rid of; nor could the career of the isolated bacilli distributed through the body be thus checked. Tuberculin has, however, found a remarkable sphere of usefulness in causing reaction in animals suffering from tuberculosis. Indeed, tuberculin is the most valuable means of diagnosis that we possess (MacFadyen). When injected (dose, 30–40 centigrammes) it causes a rise of one and a half to three degrees. The fever begins between the twelfth and fifteenth hour after injection, and lasts several hours. The duration and intensity of the reaction have no relation to the number and gravity of the lesions, but the same dose injected into healthy cattle causes no appreciable febrile reaction. The tuberculous calf reacts just as well as the adult, but the dose is generally 10–20 centigrammes. Injections of tuberculin have no troublesome effect on the quantity or quality of the milk of cows or on the progress of gestation. Cattle come first amongst animals liable to tubercle. Horses may be infected, but it is comparatively rare, and among small ruminants the disease is rarer still. Dogs, cats, and kittens may be easily infected. Amongst birds, fowls, pigeons, turkeys, and pheasants, the disease assumes almost an epidemic character. Especially do animals in confinement die of tubercle, as is illustrated in zoölogical gardens. Respecting the lesions of bovine tuberculosis, it will be sufficient to say that nothing is more variable than the localisation or form of its attacks. The lungs and lymphatic glands come first in order of frequency, next the serous membranes, then the liver and intestines, and lastly the spleen, joints, and udder (Nocard). Tuberculosis of Animals. The anatomical changes in bovine tubercle are mostly found in the lungs and their membranes, the pleuræ. It also affects the internal membrane lining, the abdomen and its chief organs, the peritoneum, and the lymphatic glands. In both these localities a characteristic condition is set up by small grey nodules appearing, which increase in size, giving an appearance of "grapes." Hence the condition is called grape disease, or . The organs, as we have said, are equally affected, and when we add the lymphatic glands we have a fairly complete summary of the form of the disease as it occurs in cattle. As has been clearly pointed out by Martin, Woodhead, and others in their evidence before the Royal Commission, the organs, glands, and membranes are the sites for tubercle, not the muscles (or "meat"). This latter is most liable to convey infection when the butcher smears it with the knife which he has used to remove tubercular organs. Perlsucht As regards the udder in its relation to milk infection, it may be desirable to state that the initial lesion, according to Nocard and Bang, takes the form of a progressive sclerosis. The interlobular connective tissue, normally scanty, becomes thickened, fibrous, and infiltrated by minute miliary granulations. The granular tissue is thus "smothered by the hypertrophy and fibrous transformation of the interstitial connective tissue" (Nocard). The walls of the ducts are thickened and infiltrated, the lumen frequently dilated by masses of yellow caseous material. On the whole it may be said that tubercle of the udder is rare. Usually only one quarter is attacked, and by preference the posterior. For some time the milk remains normal, but gradually it becomes serous and yellow, and contains coagula holding numbers of bacilli. Lastly, it becomes purulent and dries up altogether. While the milk is undergoing these changes the lesion of the udder is becoming more marked, the tissue becomes less supple, and the toughness increases almost to a wooden hardness. The general anatomical characteristics of the disease are similar to those occurring in man. The percentage of cattle suffering from tubercle varies. In Germany it appears to vary from 2 to 8 per cent. of all cattle, in Saxony 17 to 30 per cent., in England 22 per cent. approximately (in London 40 per cent.), in France 25 per cent. Lowland breeds are much more infected than mountain breeds, which possess stronger constitutions. Tuberculosis of the is less common than that of cattle, but not so rare as that of the calf (Nocard). In nine out of ten cases the pig is infected by ingestion, particularly when fed on the refuse from dairies and cheese factories. The disease follows the same course as in cattle. The finding of the bacillus is difficult, and the only safe test is inoculation (Woodhead). pig are very rarely tuberculous by nature, though there is evidence to believe that very long cohabitation with tuberculous cattle would succeed in transmitting tuberculosis to some sheep. Sheep Tuberculosis in the is relatively very rare. It attacks the organs of the abdominal cavity, especially the glands; it affects the lung secondarily as a rule. The cases are generally isolated ones, even though the animal belongs to a stud. Nocard holds that the bacillus obtained from the pulmonary variety is like the human type, whilst the abdominal variety is more like the avian bacillus. horse Nocard says : "If the dog can become tuberculous from contact with man, the converse is equally true. Infection is at any rate possible when a house-dog scatters on the floor, carpet, or bed, during its fit of coughing, virulent material, which is rendered extremely dangerous by drying, especially for children, its habitual playmates. The most elementary prudence would recommend the banishment from a room of every dog which coughs frequently, even though it only seems to be suffering from some common affection of the bronchi or lung." Tuberculosis is a common disease among the of the poultry-yard: poultry, pigeons, turkeys, pea-fowl, guinea-fowl, etc. They are infected almost exclusively through the digestive tract, generally by devouring infected secretions of previous tubercular fowls. Whatever the position or form of avian tuberculosis, the bacilli are present in enormous numbers, and are often much shorter and sometimes much longer than those met with in tuberculous mammalia, and grow outside the body at a higher temperature (43° C.). They are also said to be more resistant and of quicker growth. The species is probably identical with Koch's bacillus, though there are differences. In the nodule, which is larger than in human tuberculosis, there are few or no giant cells, and it does not so readily break down birds Nocard and others have demonstrated the fact that the of Koch is the common denominator in all tubercular disease, whatever and wherever its manifestations, in all animals. The bacillus, they hold, may, however, experience profound modifications by means of successive passages through the bodies of divers species of animals. But if the modifications which it undergoes as a result of transmissions through birds, for example, are profound enough to make the bacillus of avian tubercle a peculiar variety of Koch's bacillus, they are not enough, it is generally believed, to make these bacilli two distinct species. Bacillus tuberculosis We may, therefore, take it for granted that tuberculosis is one and the same disease, with various manifestations, common to man and animals, intercommunicable, and having but one : the of Koch. vera causa Bacillus tuberculosis At the present time much attention is being directed to the administrative personal control of tuberculosis. How greatly this is needed in so preventable a disease is evident from a perusal of the following quotation from the Registrar-General's reports. (See opposite page.) The Prevention of Tuberculosis. These figures show a marked decline in the three worst forms of the disease. But this decline is apparently less marked in tabes than in phthisis or tubercular meningitis, , less in the kind of tubercle due to the ingestion of infected milk. Fortunately the State is beginning to realise its duty in regard to preventive measures. The abolition of private slaughter-houses, the protection of meat and milk supplies, the seizure of tuberculous milch cows, and such like measures fall obviously within the jurisdiction of the State rather than the individual, and claim the earnest and urgent attention of the public health departments of states. i. e. ENGLISH DEATH-RATES FROM ALL TUBERCULAR DISEASES is tuberculosis of the alimentary canal and mesenteric lymph glands. Tabes mesenterica is the name of the same disease as it affects the membranes of the brain (acute hydrocephalus). Tubercular meningitis is the term applied to "consumption," or tubercle in the lungs. Phthisis But personal hygiene and the prevention of the transmission of the disease depend very largely indeed upon the mass of the population. Hence we hail with satisfaction the recent endeavours to educate public opinion. In order to make this matter very simple indeed, we have placed in a footnote a series of statements embodying some of the chief facts which every individual in our crowded communities should know. Bacillus of Diphtheria (Klebs-Löffler Bacillus, 1882–1884). Diphtheria is an infective disease characterised by a variety of clinical symptoms, but commonly by a severe inflammation followed by a fibrous infiltration (constituting a ) of certain parts. The membrane ultimately breaks down. The parts affected are the mucous membrane of the fauces, larynx, pharynx, trachea, and sometimes wounds and the inner wall of the stomach. The common sign of the disease is the membrane in the throat; but muscle weakness, syncope, albuminuria, post-diphtheritic paralysis, convulsions, and many other symptoms guide the physician in diagnosis and the course of the disease. Diphtheria membrane The was isolated from the many bacteria found in the membrane by Löffler. Klebs had previously identified the bacillus as the cause of the disease. It is a slender rod, straight or slightly curved, and remarkable for its beaded appearance; there are also irregular and club-shaped forms. It differs in size according to its culture medium, but is generally 3 or 4 µ in length. In the membrane which is its strictly local habitat in the body—indeed, the bacillus is found nowhere else in the body—it almost invariably shows parallel grouping, lying between the fibrin of the membrane, and most largely in its deeper parts. Here it is mixed with other bacilli, micrococci, staphylococci, and streptococci, all of which are present and performing their part in complicating the disease. The bacillus possesses five negative characters; namely, it has no spores, threads, or power of mobility, and does not produce liquefaction or gas. It stains with Löffler's methylene blue, and shows metachromatic granules and polar staining. Its favourable temperature is blood-heat, though it will grow at room temperature. It is aërobic, and, indeed, prefers a current of air. Löffler contrived a medium for cultivation which has proved most successful. It is made by mixing three parts of ox-blood serum with one part of broth containing 1 per cent. of glucose, 1 per cent. of peptone, and 1/2 per cent. of common salt; the whole is coagulated. Upon this medium the Klebs-Löffler bacillus grows rapidly in eighteen or twenty hours, producing scattered "nucleated" round white colonies, becoming yellowish. It grows well in broth, but without producing either a pellicle or turbidity; it can grow on the ordinary media, though its growth on potato is not visible; on the white of egg it flourishes extremely well. Bacillus diphtheriæ It retains its vitality in cultures and sometimes in the throat for months. Three or four weeks is the average length of time for its existence in the membrane, but, owing to the difficulty of killing it , it may live on for as long as a year. All the conditions in the throat—mucous membrane, blood-heat, moisture, air—are extremely favourable to the bacillus; but it is very materially modified in virulence. It is secured for diagnostic purposes by one of two methods: ( ) Either a piece of the membrane is detached, and after washing carefully examined by culture as well as the microscope; or ( ) a "swab" is made from the infected throat and cultured on serum, and incubated at 37° C. for eighteen hours and then microscopically examined. Both methods—and there is no further choice—present some difficulties owing to the large number of bacteria found in the throat. Hence a negative result must be accepted with reserve. in situ a b We have already referred at some length to the question of in diphtheria, and need not dwell further upon that matter. Still a word or two may be said here summarising the general action of the bacillus. Locally it produces inflammatory change with fibrinous exudation and some cellular necrosis. In the membrane a ferment is probably produced which, unlike the localised bacilli, passes throughout the body and by digestion of the proteids produces albumoses and an organic acid which have the toxic influence. The toxins act on the blood-vessels, and nerves, and muscle fibres of the heart, and many of the more highly specialised cells of the body. Thus we get degenerative changes in the kidney, in cells of the central nervous system, in the peripheral nerves (post-diphtheritic paralysis), and elsewhere, these pathological conditions setting up, in addition to the membrane, the signs of the disease. The bacil lus is pathogenic for the horse, ox, rabbit, guinea-pig, cat, and some birds. Cases are on record of supposed infection of children by cats suffering from the disease. The horse, it will be remembered, yields the antitoxin which has saved so many lives (Metropolitan Asylums Board Report, 1896). toxins The influence of drainage, milk, and schools must not be forgotten by sanitary authorities any more than the essential importance of adequate isolation hospital accommodation. Mr. Shattock's experiments on the effect of sewer air upon attenuated Klebs-Löffler bacilli have been mentioned (see p. 105). Nevertheless there can be no doubt that emanations from defective drains have a materially predisposing effect, not, it is true, upon the bacilli, but upon the tissues. Sore throats thus acquired are the site for the development of diphtheria. par excellence The influence of school attendance has claimed the recent attention of the Medical Officer of the London School Board and the Medical Officer of the administrative County of London. In London since 1881 there has been a marked increase of diphtheria, which has occurred, though in a much less degree, throughout England and Wales. The Registrar-General has only classified diphtheria as a separate disease since 1855, when the death-rate per 1,000,000 in England and Wales was stated as 20. The following are the figures for four decades up to 1895: average death-rate per million of the population from diphtheria in england and wales and in london (in decades 1856–95) From these figures the extraordinary increase during the last few years is clearly demonstrated. Sir Richard Thorne Thorne, in 1891, drew attention to the influence of damp soils and schools upon diphtheria. In 1894 Mr. Shirley Murphy, Medical Officer to the London County Council, reported that there had been an increase in diphtheria mortality in London at school ages (three to ten) as compared with other ages since the Elementary Education Act became operative in 1871; that the increased mortality from diphtheria in populous districts, as compared with rural districts, since 1871, might be due to the greater effect of the Education Act in the former; and that there was a diminution of diphtheria in London during the summer holidays at the schools in 1893, but that 1892 did not show any marked changes for August. In 1896 Professor W. R. Smith, the Medical Officer to the London School Board, furnished a report on this same subject of school influence, in which he produces evidence to show that the recrudescence of the disease in 1881–90 was greatest in England and Wales at the age of two to three years, and in London at the age of one to two years, in both cases ; that age as an absolute factor in the incidence of the disease is enormously more active than any school influence, and that personal contact is another important source of infection. before school age Although it is said that "statistics can be made to prove anything," there can be little doubt that both of these reports contain a great deal of truth; nor are these truths incompatible with each other. They both emphasise as a great factor in the incidence of the disease, and whatever affects the health of the child population, like schools, must play, directly or indirectly, a not unimportant part in the transmission of the disease. age Löffler and Hoffman described a bacillus having the same morphological characters as the true , except that it had no virulence. Roux believes this is merely an attenuated diphtheria bacillus. It is frequently found in healthy throats. The chief differences between the real and the pseudo-bacillus are: The Pseudo-diphtheria Bacillus. Bacillus diphtheriæ 1. The pseudo-bacillus is thicker in the middle than at the poles, and not so variable as the . Polar staining is absent. Bacillus diphtheriæ 2. Its growth on potato reveals cream-coloured colonies visible in a couple of days; the real bacillus is invisible. 3. The pseudo-bacillus will not grow at all anaërobically in hydrogen, but the is able to do so. Bacillus diphtheriæ 4. There is the great difference in virulence. Suppuration. This term is used to designate that general breaking down of cells which follows acute inflammation. An "abscess" or "gathering" is a collection, greater or smaller, of the products of suppuration. The word is generally used to describe this matter. We may have such an advanced inflammatory condition in any locality of the body, and it will assume different characters according to its site. Hence there are connected with suppuration, as causal agents, a variety of bacteria. Pus is not matter containing a pure culture of any specific species, but, on the contrary, is generally filled with a large number of different species. The most important are as follows: pus 1. These are micrococci arranged in groups, which have been likened to bunches of grapes. They are the common organisms found in pus, and were with other auxiliary bacteria first distinguished as such by Professor Ogston, of Aberdeen. There are several forms of the same species, differing from each other in colour Staphylococcus pyogenes aureus. Thus we have the (golden yellow), (white), (lemon), and others. They occur commonly in nature, in air, soil, water, on the surface of the skin, and in all suppurative conditions. The is the only one credited with much virulence. It occurs in the blood in blood-poisoning (septicæmia, pyæmia), and is present in all ulcerative conditions, including ulcerative disease of the valves of the heart. S. pyogenes aureus albus citreus aureus The and are slightly modified forms of the , and are differentiated from it by being non-liquefying. They produce a wax-like growth on gelatine. Staphylococcus cereus albus S. cereus flavus S. pyogenes aureus , the type of the family, is grown in all ordinary media at room temperature, though more rapidly at 37° C. Liquefaction sets in at a comparatively early date, and subsequently we have in the gelatine test-tube cultures a flocculent deposit of a bright yellow amorphous mass, and in gelatine plates small depressions of liquefaction with a yellow deposit. It renders all media acid, and coagulates milk. Its thermal death-point in gelatine is 58° C. for ten minutes, but when dry considerably higher. It is a non-motile and a facultative anaërobe; but the presence of oxygen is necessary for a bright colour. Its virulence readily declines. Staphylococcus pyogenes aureus 2. In this species of micrococcus the elements are arranged in chains. Most of the streptococci in pus, from different sources, are one species, having approximately the same morphological and biological characters. Their different effects are due to different degrees of toxic virulence; they are always more virulent when associated with other bacteria, for example, the Proteus family. Streptococcus pyogenes. The chains vary in length, consisting of more elements when cultured in fluid media. They multiply by direct division of the individual elements, and in old cultures it has been observed that the cocci vary in form and size. This latter fact gave support to the theory that streptococcus reproduced itself by , or "mother-cells." arthrospores In culture upon the ordinary media streptococcus is comparatively slow-growing, producing minute white colonies on or about the sixth day. It does not liquefy gelatine, and remains strictly localised to the track of the inoculating needle. Like the staphylococcus, it readily loses virulence. The thermal death-point is, however, lower: 54° C. for ten minutes. Marmorek has devised a method by which the virulence may be greatly increased, and he holds that it depends upon the degree of virulence possessed by any particular streptococcus as to what effects it will produce. By the adoption of Marmorek's methods attempts have been made to prepare an antitoxin. has been isolated from the membrane of diphtheria, and from small-pox, scarlet fever, vaccinia, and other diseases. In such cases it is not the causal agent, but merely associated with the complications of these diseases. Suppuration and erysipelas are the only pathological conditions in which the causal agency of streptococcus has been sufficiently established. Streptococcus pyogenes 3. The occurs in green pus, and is the cause of that colouration. Gessard was the first to prove its significance, and he describes two varieties. Bacillus pyocyaneus Micrococcus Tetragonus It is a minute, actively motile, non-sporulating bacillus, which occasionally complicates suppuration and produces green pus. Oxygen is necessary for pigmentation, which is due to two substances: , a greenish-blue product extracted with chloroform, and , a brown substance derived from the oxidation of the former pigment. Both these colours are produced in cultivation outside the body. On gelatine the colour is green, passing on to olive. There is liquefaction. On potato we generally obtain a brown growth (compare , , and others). The organism grows rapidly on all the ordinary media, which it has a tendency to colour throughout. pyocyanin pyoxanthose Bacillus coli B. mallei It will be remembered that when speaking of the antagonism of organisms, we referred to the inimical action of upon anthrax. Bacillus pyocyaneus 4. This species occurs in phthisi cal cavities and in certain suppurations in the region of the face. It is a micrococcus usually in the form of small tetrads. A capsule is always present and sometimes discernible. Micrococcus Tetragonus. 5. and many putrefactive germs commonly occur in suppurative conditions, but they are not restricted to such disorders (see p. 64). Bacillus coli communis Diplococcus of Neisser 6. (Neisser, 1879). This organism is more frequently spoken of as a diplococcus. It occurs at the acute stage of the disease, but is not readily differentiated from other similar diplococci except by technical laboratory methods. Each element presents a straight or concave surface to its fellow. A very marked concavity indicates commencing fission. The position which these diplococci take up in pus is , and arranged more or less definitely around the nucleus. Difficulty has often been found in cultivating this organism in artificial media outside the body. Wertheim and others have suggested special formulæ for the preparation of suitable media, but it is a very simple matter to secure cultures on agar plates smeared with human blood from a pricked finger. The plate is incubated at 37° C. At the end of twenty-four hours small raised grey colonies appear, which at the end of about four days show adult growth. The margin is uneven, and the centre more opaque than the rest of the colony. This diplococcus is readily killed, and sub-cultures must be frequently made to retain vitality and virulence. Light, desiccation, and a temperature of 55° C. all act germicidally. The organism stains readily in Löffler's blue, but is decolourised by Gram's method. It is more or less strictly parasitic to man. Its shape, size, character of growth, and staining properties assist in differentiating it from various similar diplococci. Micrococcus gonorrhœæ intracellular Anthrax. This disease was one of the first in which the causal agency of bacteria was proved. In 1849 Pollender found an innumerable number of small rods in the blood of animals suffering from anthrax. In 1863 Davaine described these, and attributed the disease to them. But it was not till 1876 that Koch finally settled the matter by isolating the bacilli in pure culture and describing their biological characters. It is owing in part to its interesting bacterial history, which opened up so much new ground in this comparatively new science, that anthrax has assumed such an important place in pathology. But for other reasons, too, it claims attention. It appears to have been known in the time of Moses, and was perhaps the disease described by Homer in the First Book of the Iliad. Rome was visited by it in 740 B.C. Anthrax is an acute disease, affecting sheep, cattle, horses, goats, deer, and man. Cats, white rats, and Algerian sheep are immune. Swine become infected by feeding on the offal of diseased cattle (Crookshank). The signs are mainly three: The is greatly enlarged and congested, is friable to the touch, and contains enormous numbers of bacilli; the may show exudations forming dark gelatinous tumours; and the remains fluid for some time after death, is black, tar-like, contains bubbles of air, and shows other degenerative changes in the red corpuscles, whilst the contain such vast quantities of bacilli that they may be ruptured by them. Particularly is this true in the peripheral arteries. Many of the organs of the body show marked congestion. post-mortem spleen skin blood small blood-vessels Clinically there is rise of temperature and rapid loss of muscular power. The of anthrax are square-ended rods 1 µ broad and 4–5 µ long. In the tissues of the body they follow the lines of the capillaries, and are irregularly situated. In places they are so densely packed as to form obstructions to the onward flow of blood. In cultures they are in chains end to end, having as a rule equal interbacillary spaces. In cultures long filaments and threads occur. The exact shape of the bacillus depends, however, upon two things: the staining and spore formation. Both these factors may very materially modify the normal shape. The spores of anthrax are oval endospores, produced only in the presence of free oxygen, and at any temperature between 18 and 41° C. On account of requiring free oxygen, they are formed only outside the body. The homogeneous pro toplasm of the bacillus becomes granular; the granules coalesce, and we have spores. Each spore possesses a thick capsule, which enables it to resist many physical conditions which kill the bacillus. When the spore is ripe or has exhausted the parent bacillus, it may take on a resting stage, or under favourable circumstances commence germination, very much after the manner of a seed. The spores may infect a farm for many months; indeed, cases are on record which appear to prove that the disease on a farm in the autumn may by means of the spores be carried on by the hay of the following summer into a second winter. Thus, by means of the spores, the infection of anthrax may cling to the land for very long periods, even for years. Spores of anthrax can withstand 5 per cent. carbolic acid or 1–1000 corrosive sublimate for more than an hour; even boiling does not kill them at once, whilst the bacilli without their spores are killed at 54° C. in ten minutes. When the spores are dry they are much more resistant than when moist. Hence the persistence of the anthrax bacillus is due to its spores. bacilli The bacillus is aërobic, non-motile, and liquefying. Broth cultures become turbid in thirty-six hours, with nebulous masses of threads matted together. The pellicle which forms on the surface affords an ideal place for spore formation. Cultures in the depth of gelatine show a most characteristic growth. From the line of inoculation delicate threads and fibrillæ extend outwards horizontally into the medium. Liquefaction commences at the top, and eventually extends throughout the tube. On gelatine plates small colonies appear in thirty-six hours, and on the second or third day they look, under a low power of the microscope, like matted hair. The colonies after a time sink in the gelatine, owing to liquefaction. On potato, agar, and blood serum anthrax grows well. 1. This is the usual mode of infection in animals grazing on infected pasture land. A soil suitable for the propagation of anthrax is one containing abundance of air and proteid material. Feeding on bacilli alone would probably not produce the disease, owing to the germicidal effect of the gastric juice. But spores can readily pass uninjured through the stomach and produce anthrax in the blood. Infected water as well as fodder may convey the disease. Water becomes infected by bodies of animals dead of anthrax, or, as was the case once at least in the south-west of England, by a stream passing through the washing-yard of an infected tannery. Manure on fields, litter in stalls, and infected earth may all contribute to the transmission of the disease. Darwin pointed out the services which are performed in superficial soils by earthworms bringing up casts; Pasteur was of opinion that in this way earthworms were responsible for continually bringing up the spores of anthrax from buried corpses to the surface, where they would reinfect cattle. Koch disputed this, but more recently Bollinger has demonstrated the correctness of Pasteur's views by isolating anthrax contagium from five per cent. of the worms sent him from an anthrax pasture. Bollinger also maintains that flies and other insects may convey the disease from discharges or carcasses round which they congregate. Channels of Infection. The Alimentary Canal. Alimentary infection in man is a rare form, and it reveals itself in a primary diseased state known as , an inflamed condition of the intestine and mesenteric lymph glands. mycosis intestinalis 2. Cutaneous anthrax goes by the name of , and is caused by infective anthrax matter gaining entrance through abrasions or ulcers in the skin. This local form is obviously most contracted by those whose occupation leads them to handle hides or other anthrax material (butchers and cleaners of hides). Two or three days after inoculation a red pimple appears, which rapidly passes through a vesicular stage until it is a pustule. Concomitantly we have glandular enlargement, general malaise, and a high temperature. Thus from a local sore a general infection may result. Unless this does occur, the issue will not be fatal, and the bacilli will never gain entrance into the blood or be anything but local. Through the Skin. malignant pustule 3. In man this is the commonest form of all, and is well known under the term "wool-sorters' disease," or pulmonary anthrax. This mode of infection occurs when dried spores are inhaled in processes of skin-cleaning. It frequently commences as a local lesion affecting the mucous membrane of the trachea or bronchi, but it rapidly spreads, affecting the neighbouring glands, which become greatly enlarged, and extending to the pleura and lung itself. Such cases, as a rule, rapidly end fatally. Respiratory Tract. From what has been said, it will be clear that anthrax carcasses are better not opened and exposed to free oxygen. An extended examination is not necessary. Burning the entire carcass in a crematorium would be the ideal treatment. As such is not generally feasible the next best thing is to bury the carcass deeply with lime below and above it, and rail in the area to prevent other animals grazing off it. post-mortem A very small prick will extract enough blood to examine for the anthrax bacilli which are driven by the force of the blood-current to the small surface capillaries. This occurs, of course, only when the disease has become quite general, for in the early stage the healthy blood limits the bacilli to the internal organs. In such cases examination of the blood of the spleen is necessary. Anthrax covers a wide geographical area all over the world, and no country seems altogether exempt. In Germany as many as 3700 animals have been lost in a single year. About 900 animals were attacked in 1897 in Great Britain. Plague. This disease, like anthrax and leprosy, has a long historical record behind it. As the Black Death it decimated the population of England in the fourteenth century, and visited the country again in epidemic form in the middle of the seventeenth century, when it was called the Great Plague. Now, it is highly probable that these two scourges and the recent epidemic in the East are all forms of one and the same disease. As a matter of fact, it is difficult to be sure what was the exact pathology of a number of the grievous ailments which troubled our country in the Middle Ages, but from all accounts bubonic plague and true leprosy were amongst them. The former came and went spasmodically, as is its habit; the latter dragged through the length of several centuries. Bacillus of Plague The distribution of plague at the present time is fortunately a somewhat limited one, namely, a definite area in Asia known as the "Plague Belt." From Mesopotamia, as a sort of focus, the disease spreads northwards to the Caspian Sea, westwards to the Red Sea, southwards as far as Central India, and eastwards as far as the China Sea. This constitutes the "belt," but the disease may take an epidemic form, and is readily, though very slowly, conveyed by infection or contagion. It appears to be infectious by means of infective dust, and contagious by prolonged and intimate contact with the plague-stricken. Rats have been accused of conveying the disease from port to port, and even infecting man. It is clear that rats are not the only agency acting in this way. Nevertheless it is true that rats contract the disease more readily than any other animals, and that when suffering from it they may spread the infection. How it is thus spread it is not known. Drs. Cantlie and Yersin have pointed out that previously to an epidemic of plague rats die in enormous numbers. The bacteriology of plague is almost the latest addition to the science. Kitasato, of Tokio, demonstrated the cause of plague to be a bacillus during the Hong Kong epidemic in 1894. This was immediately confirmed by Yersin, and further proved by the isolation in artificial media of a pure culture of a bacillus able to cause the specific disease of bubonic plague. The bacillus was first detected in the blood of patients suffering from the disease. It takes the form of a small, round-ended, oval cell, with marked polar staining, and hence having an appearance not unlike a diplococcus. In the middle there is a clear interspace, and the whole is surrounded with a thick capsule, stained only with difficulty. The organisms are often linked together in pairs or even chains, and exhibit involution forms. In culture the bacillus is even more coccal in form than in the body. The plague bacillus grows readily on the ordinary media at blood-heat, producing circular cream-coloured colonies with a wavy outline, which eventually coalesce to form a greyish film. The following negative characters help to distinguish it: No growth occurs on potato, milk is not coagulated, and gelatine is not liquefied; Gram's method does not stain the bacillus, and there are no spores; the bacillus is readily killed by heat and by desiccation over sulphuric acid at 30° C. Both in cultures and outside the body the bacillus loses virulence. To this may be attributed possibly the variety of forms of the plague bacillus which differ in virulence. On gaining entrance to the human body the bacillus affects in particular two organs, the spleen and the lymph glands. The latter become inflamed in groups, commencing frequently with those in the armpit (axillary) or groin (inguinal). The spleen suffers from inflammatory swelling, which may affect other organs also. In both places the bacilli occur in enormous numbers. Kitasato considers that the bacillus may enter the body by the three channels adopted by anthrax, namely, the skin, alimentary canal, and respiratory tract. Haffkine has prepared a vaccine to be used as a prophylactic. He grows a pure culture of Kitasato's bacillus in broth upon the surface of which some globules of fat ("ghee") have been placed. The bacillus grows upon this fat in copious stalactite form. From time to time this growth is shaken down, until after five or six weeks the shaken broth appears milky. The contained bacilli are killed by heating the fluid to 70° C. for one hour. The resultant is the vaccine, of which the dose is 3 cc. Haffkine believes that inoculated persons in India have suffered twenty times less than non-inoculated living under the same conditions. Plague is essentially a "filth disease," and it is frequently preceded by famine. Temperature and overcrowding exert an influence upon it. The areas affected by the disease in the Middle Ages, in the seventeenth century, and in 1894–96 are alike in being characterised by filth and overcrowding. There is little fear, speaking generally, of the plague ever flourishing under Western civilisation, where the conditions are such that even when it appears there is little to encourage or favour its development. Leprosy. This ancient disease is said to have existed in Egypt 3500 B.C., and was comparatively common in India, China, and even in parts of Europe 500 B.C. We know it has existed in many parts of the world in the past, in which regions it is now extinct. Some of the earliest notices we have of it in this country come from Ireland, and date back to the fifth and sixth centuries. Even at that period of time also various classical descriptions of the disease had been written and various decrees made by the Church councils to protect lepers and prevent the spread of the disease, which was often looked upon as a divine visitation. In the tenth century leprosy was prevalent in England; it reached its zenith in the thirteenth century, or possibly a little earlier, and declined from that date to its extinction in the sixteenth. But even two hundred years later leprosy was endemic in the Shetlands, and it is recorded that in 1742 there was held a public thanksgiving in Shetland on account of the disappearance of leprosy. At one time or another there were as many as two hundred institutions in the British Isles for the more or less exclusive use of lepers. Many of these establishments were of an ecclesiastical or municipal character, and probably the exact diagnosis of diseases was a somewhat lax matter. Bury St. Edmunds, Bristol, Canterbury, London, Lynn, Norwich, Thetford, and York were centres for lepers. Burton Lazars and Sherburn, in Durham, were two of the more famous leper institutions. At the present time the distribution of the disease is mostly Asiatic. Norway contains about 1200 lepers, Spain approximately the same number. Scattered through Europe are perhaps another 600 or 700, in India 100,000, and a large number in Japan. The Cape possesses a famous leper hospital on Robben Island, with a number of patients. The disease is also endemic in the Sandwich Islands. Descriptions of the pathological varieties of leprosy have been very diverse. The classification now generally adopted includes three forms: the , the (or maculo-anæsthetic), and the . is that form of the disease affecting chiefly the skin, and resulting in nodular tuberculated growth or a diffuse infil tration. It causes great disfigurement. The anæsthetic form causes a destruction of the nerve fibres, and so produces anæsthesia, paralysis, and what are called "trophic" changes. Not infrequently patches occur on the skin, which appear like parchment, owing to this trophic change. Bullæ may arise. When the tissue change is radical or far advanced, considerable distortion may result. The mixed variety of leprosy, as its name implies, is a mixture of the two other forms. tuberculated anæsthetic mixed Lepra tuberculosa The was discovered by Hansen in 1874. He found it in the lepra cells in the skin, lymph glands, liver, spleen, and thickened parts of the nerves. It is common in the discharges from the wounds of lepers. It is conveyed in the body by the lymph stream, and has rarely been isolated from the blood (Köbner). Bacillus lepræ The bacillus is present in enormous numbers in the skin and tissues, and has a form very similar indeed to . It is a straight rod, and showing with some staining methods marked beading, but with others no beading at all. It measures 4 µ long and 1 µ broad. Young leprosy bacilli are said to be motile, but old ones are not. Neisser has maintained that the bacillus possesses a capsule and spores. The latter have not been seen, but Neisser holds that this is the form in which the bacillus gains entrance to the body. There is a characteristic which fortunately aids us in the diagnosis of this disease in the tissues, and that is the arrangement of the bacilli, which are rarely scattered or isolated, but gathered together in clumps and colonies. Bordoni-Uffreduzzi and Campania claim to have isolated the bacillus and grown it on artificial media, the former aërobically on peptone-glycerine-blood-serum, at 37° C., the latter anaërobically. But no other worker has been able to do this. Hence we are not able to study the bacteriology of leprosy at all completely, nor have inoculation experiments proved successful. Nevertheless there is little doubt that leprosy is a bacterial disease produced by the bacillus of Hansen. Bordoni-Uffreduzzi maintains that the parasitic existence of the may alternate with a saprophytic stage. This may be of importance in the spread of the disease. There is evidence in support of the non-communicability of the disease by heredity or contagion. Segregation does not appear always to result in a decline of the disease, as we should expect if it were purely contagious. Ehlers, of Copenhagen, has, however, as recently as 1897, reaffirmed his belief in the contagiousness of leprosy; Virchow, on the other hand, has declared that it is not highly contagious. There is evidence to show that persons far advanced in the disease may live in a healthy community and yet not infect their immediate neighbours. Indeed, the transmission of the disease is still an unsolved problem. Mr. Hutchinson suggests diet, particularly uncooked or putrid fish, as a likely channel; on the other hand, leprosy appears in districts where no fish is eaten. Deficiency of salt, telluric and climatic conditions, racial tendencies, social status, poverty, insanitation, drinking water, even vaccination, have all secured support from various seekers after the true channel by which the bacillus gains entrance to the human body. The real mode of transmission is, however, still unknown. The decline and final extinction of leprosy in the British Islands was probably due in part to the natural tendency of the disease, under favourable hygienic circumstances, to die out, and in part to a general and extensive social improvement in the life of the people, to a complete change in the poor and insufficient diet, and to agricultural advancement, improved sanitation, and land drainage. Bacillus tuberculosis Bacillus lepræ At the Leprosy Congress held in Berlin in 1897, Hansen again emphasised his belief that segregation was the cause of the decline of leprosy wherever it had occurred. But there appears to be some evidence to show that leprosy has declined where there has been no segregation whatever, and therefore, however favourable to decline such isolation may be, it would seem not to be actually necessary to the decline. At the same Congress Besnier declared in favour of the infective virus being widely propagated by means of the nasal secretion. Sticker states that the nasal secretion contains millions of lepra bacilli, especially in the acute stage of the disease, and Besnier and Sticker have pointed out how frequently and severely the septum nasi and skin over the nose are affected in leprosy. Several leprologists in India have recorded similar observations. These facts appear to support Besnier's contention that the disease is spread by nasal secretion. We may fitly add here the conclusions arrived at by the English Leprosy Commission in India: "1. Leprosy is a disease ; it is not a form of syphilis or tuberculosis, but has striking etiological analogies with the latter. sui generis "2. Leprosy is not diffused by hereditary transmission, and, for this reason and the established amount of sterility among lepers, the disease has a natural tendency to die out. "3. Though in a scientific classification of diseases leprosy must be regarded as contagious, and also inoculable, yet the extent to which it is propagated by these means is exceedingly small. "4. Leprosy is not directly originated by the use of any particular article of food, nor by any climatic or telluric conditions, nor by insanitary surroundings, neither does it peculiarly affect any race or caste. "5. Leprosy is indirectly influenced by insanitary surroundings, such as poverty, bad food, or deficient drainage or ventilation, for these by causing a predisposition increase the susceptibility of the individual to the disease. "6. Leprosy, in the great majority of cases, originates , that is, from a sequence or concurrence of causes and conditions dealt with in the Report, and which are related to each other in ways at present imperfectly known." de novo The practical suggestions of the Commission for preventive treatment included voluntary isolation, prohibition of the sale of articles of food by lepers, leper farms, orphanages, and "improved sanitation and good dietetic conditions" generally. Serum-therapy has been attempted on behalf of the French Academy of Medicine, but without success. Many forms of treatment ameliorate the miserable condition of the leper, but up to the present no curative agent has been found. Diplococcus of Pneumonia Pneumonia. Some of the difficulty which has surrounded the bacteriology of inflammation of the lungs is due to the confusion arising from supposing that attacks of the disease differed only in degree. Pneumonia, however, has various forms, arising now from one cause, now from another. The specific or croupous pneumonia is associated with two organisms: Fraenkel's diplococcus and Friedländer's pneumo-bacillus. Several other bacteria have from time to time been held responsible for pneumonia, a streptococcus re ceiving, at one time, some support. But whilst opinion is divided on the rôle of various extraneous and concomitant bacteria in lung disease, importance is attached to Fraenkel's and Friedländer's organisms. The of Fraenkel is a small, oval diplococcus found in the "rusty" sputum of croupous pneumonia. It is non-motile, non-liquefying, and aërobic. When examined from cultures the diplococci are frequently seen in chains, not unlike a streptococcus, and there is some reason to suppose that this form gave rise to the belief that it was another species; when examined from the tissues it possesses a capsule, but in culture this is lost. It is difficult to cultivate, but grows on glycerine agar and blood serum at blood-heat. On ordinary gelatine at room temperature it does not grow, or, if so, very slightly. The ideal fluid is a slightly alkaline liquid medium, and in twenty-four hours a powdery growth will occur in such broth. On potato there is apparently no growth. It rapidly loses its virulence on solid media, and is said to be non-virulent after three or four sub-culturings. A temperature of 54–58° C. for a few minutes kills the bacteria, but not the toxin. This, however, is removed by filtration, and is therefore probably intracellular. It is attenuated by heating to 70° C. diplococcus Fraenkel's diplococcus occurs, then, in the acute stage of pneumonia, in company with streptococci and staphylococci. It also occurs in the blood in certain suppurative conditions, in pleurisy and inflammation of the pericardium, and sometimes in diphtheria, and therefore it is not peculiar to pneumonia. There is one other point to which attention should be drawn. Fraenkel's organism is said to be frequently present in the saliva of healthy persons. Pneumonia depresses the resistant vitality of the tissues, and thus affords to the diplococcus present in the saliva an excellent nidus for its growth. is a capsulated oval coccus, assuming the form of a small bacillus. It is inconstant in pneumonia, unequally distributed, and scarce; it is aërobic, and facultatively anaërobic; it occasionally occurs in long forms and filaments; it is non-motile, non-liquefying, and has no spores; it does not stain by Gram's method, which stain is therefore used for differential diagnosis; it will grow fairly well in ordinary gelatine at 20° C.; and it is a denitrifying organism, and also an actively fermentative one, even fermenting glycerine. It is not unlike , and to distinguish it from that organism we may remember that the is motile, never has a capsule, produces indol, and does not ferment glycerine. Friedländer's Pneumo-bacillus Bacillus coli communis B. coli Bacillus of Influenza Influenza. In 1892, during the pandemic of influenza, Pfeiffer discovered a bacillus in the bronchial mucus of patients suffering from the disease. It is one of the smallest bacilli known, and frequently occurs in chains not unlike a streptococcus. Carron obtained the same organism from the blood. In the bronchial expectoration it can retain its virulence for as long as a fortnight, but it is quickly destroyed by drying. The bacillus is aërobic, non-motile, and up to the present spores have not been found. It grows somewhat feebly in artificial media, and readily dies out. Blood serum, glycerine agar, broth, and gelatine have all been used at blood-heat. It does not grow at room tem perature. Pfeiffer's bacillus appears most abundantly at the height of the disease, and disappears with convalescence. It is said not to appear in any other disease. Yellow Fever. Sternberg and Havelburg have both isolated bacilli from cases of yellow fever; but the organism discovered by Sanarelli, the , is now accepted as the causal agent of the disease. It is a small, short rod, with round ends, and generally united in pairs; it has various pleomorphic forms; it grows well on all the ordinary media; it is killed in sea-water at 60° C., and also by direct sunlight in a few hours. Bacillus icteroides Diarrhœa of Infants. From time to time different organisms have been isolated in this diseased condition. and (Klein) have been held responsible for it. W. D. Booker, of Johns Hopkins University, sums up an extended research into the question as follows: Bacillus coli B. enteriditis sporogenes "No single micro-organism is found to be the specific exciter of the summer diarrhœa of infants, but the affection is generally to be attributed to the result of the activity of a number of varieties of bacteria, some of which belong to well-known species, and are of ordinary occurrence and wide distribution, the most important being the streptococcus and . Proteus vulgaris "The first step in the pathological process is probably an injury to the epithelium from abnormal or excessive fermentation, from toxic products of bacteria, or from other factors. "Bacteria exert a direct injury upon the tissues in some instances, whereas in others the damage is brought about indirectly through the production of soluble poisons." Actinomycosis. This disease affects both animals and man. As Professor Crookshank points out, it has long been known in this country, but its various manifestations have been mistaken for other diseases or have received popular names. Here we can only mention the most outstanding facts concerning the disease. It is caused by the "ray fungus," or , which, growing on certain cereals, often gains entrance to the tissues of man and beast by lacerations of the mucous membrane of the mouth, by wounds, or by decayed teeth. Barley has been the cereal in question in some cases. The result of the introduction of the parasite is what is termed an "infective granuloma." This is, generally speaking, of the nature of an inflammatory tumour composed of round cells, epithelioid cells, giant cells, and fibrous tissue, forming nodules of varying sizes. In some cases they develop to large tumours, in others they soon break down. Actinomycosis in some ways closely resembles tuberculosis in its tissue characters. Streptothrix actinomyces In the discharge or pus from human cases of the disease small sulphur-yellow bodies may be detected, and these are tufts of "_clubs_" which are the broken-down rays of the parasite; for in the tissues which are affected the parasite arranges itself in a radiate manner, growing and extending at its outer margin and degenerating behind. In cattle the centre of the old ray becomes caseated, like cheese, or even calcified, like a stone. In the human disease abundant "_threads_" are formed as a tangled mass in the middle of the colony. As clubs characterise the bovine actinomycosis, so threads are a feature of the human form of the disease. But in both there is a third element, namely, , called by some spores, by others simply cocci. Authorities are not yet agreed as to the precise significance and rôle of these round cells. The life-history of the micro-organism may be summed up thus: small round cells "The spores sprout into excessively fine, straight or sinuous, and sometimes distinctly spirilliform threads, which branch irreg ularly and sometimes dichotomously. The extremities of the branches develop the club-shaped bodies. The clubs are closely packed together, so that a more or less globular body is formed, with a central core composed of a dense mass of threads" (Crookshank). Possibly these clubs represent organs of fructification, and produce the spores. These latter are, it is believed, set free in the vicinity of the ray, and create fresh centres of disease. In the disease manifests itself in various parts according to the locality of entrance. When occurring in the mouth it attacks the lower jaw most frequently. In one recorded case the disease was localised to the bronchi, and did not even extend into the lungs. It was probably contracted by inhalation of the parasite. The disease may spread to distant parts by means of the blood stream, and frequently the abscesses are apt to burrow in various directions. man In the the disease remains much more localised, and frequently occurs in the lower jaw, palate, or tongue. In the last site it is known as "wooden tongue," owing to the hardness resulting. The skin and subcutaneous tissues are also a favourite seat of the disease, producing the so-called wens or clyers so commonly seen in the fen country (Crookshank). Actinomycosis in cattle is specially prevalent in river valleys, marshes, and on land reclaimed from the sea. The disease occurs at all seasons, but perhaps more commonly in autumn and winter. It is more frequently met with in young animals. The disease is probably not hereditary nor readily communicated from animal to animal. ox Actinomyces may be cultivated, like other parasitic diseases, outside the body. Gelatine, blood serum, agar, glycerine agar, and potato have been used for this purpose. After a few days on glycerine agar at the temperature of the blood little white shining colonies appear, which in crease and coalesce. In about ten days' time the culture often turns a bright yellow, though it may remain white or even take on a brown or olive tint. The entire mass of growth is raised dry and crinkled, and composed almost exclusively of threads. In its early stage small bacillary forms occur, and in its later stage coccal forms. True clubs never occur in pure cultures, although the threads may occasionally show bulbous endings. Glanders in the horse and ass, and sometimes by communication in man also, is caused by a short, non-motile, aërobic bacillus, named, after the old Roman nomenclature ( ), . It was discovered in 1882 by Löffler and Schütz. It is found in the nasal discharge of glandered animals. In appearance the bacillus is not unlike , except that it is shorter and thicker. The beading of the bacillus of glanders, like that in tubercle, does not denote spores. can be cultivated on the usual media, especially on glycerine agar and potato. On the latter medium it forms a very characteristic honey-like growth, which later becomes reddish-brown. malleus Bacillus mallei B. tuberculosis B. mallei In the horse glanders particularly affects the nasal mucous membrane, forming nodules which degenerate and emit an offensive discharge. From the nose, or nasal septum, as a centre, the disease spreads to surrounding parts. It may also occur as nodules in and under the skin, when it is known as "farcy." Persons attending a glandered animal may contract the disease, often by direct inoculation. is a substance analogous to tuberculin, and is made by growing a pure culture of in glycerine-veal broth in flat flasks, with free access of calcined air. After a month's growth the culture is sterilised, filtered, concentrated, and mixed with an equal volume of a .5 per cent. solution of carbolic. The dose is 1 cc., and it is used, like tuberculin, for diagnostic purposes. If the suspected animal reacts to the injection, it is suffering from glanders. Reaction is judged by three signs, namely, a rise of temperature 2–3° C., a large "soup-plate" swelling at the site of inoculation, and an enlargement of the lymphatic glands. Mallein Bacillus mallei , , , , , and other diseases of animals have micro-organisms intimately related to them. Swine fever foot-and-mouth disease chicken cholera dysentery rinderpest There is a group of diseases due to the presence in the blood or tissues of , that is, protozoa which can live and perform their function in the blood. Amongst these are malaria, sleeping sickness, and other tropical diseases in man, and surra and various hæmatozoa in horses, fish, frogs, or rats. hæmatozoa Although a has been described as the cause of this disease, it is now almost universally supposed that the true cause is a protozoan parasite. In 1880 Laveran first described this organism, and the discovery was confirmed by Marchiafava, Celli, and others. Laveran claimed that it occurred in four different forms during the progress of its life-history: Malaria. Bacillus malariæ ( ) attached to the blood corpuscle, or free in the blood plasma. They are a little smaller than the blood-cells, and may or may not contain pigment. They eventually invade the corpuscles, possess more pigment, and lose their amœboid movement. Within the red blood corpuscles they increase in size until they reach the adult stage. a Spherical or Irregular Bodies ( ) , often assuming a rosette shape, follow next. They are pigmented, are possibly a sporing stage, and are finally set free in the blood. b Segmentation Forms ( ) , or , are free in the blood, but motionless. They are colourless, have a distinct membrane, and generally show a little pigment about the middle; they taper towards the poles. They appear in the blood after the fever has existed for some time, occurring chiefly, sometimes only, in the quotidian and malignant types of malaria. c The Crescents Semilunar Bodies ( ) apparently occur only in the blood outside the body. They are extracorpuscular bodies, and possess several long flagella, and are therefore actively motile. They are derived from the crescents or irregular intracorpuscular bodies. d The Flagellated Bodies What is the precise significance of these various forms and modifications of them is not at present known. Possibly the semilune is a resting stage inside the body, and the flagellated body another similar stage outside. Attempts to cultivate the parasite outside the body have failed. There is a good deal of evidence to show that the mosquito is the host outside the human body. There may be different forms and varieties of parasite, if not actually different species, causing the diverse forms of clinical malaria. The above account of diseases caused by bacteria does not profess to be in any sense exhaustive. It is merely illustrative. It reveals some of the disease-producing powers of micro-organisms. There are a large number of other diseases in which bacteria have been found. They are not the causes, but only accidentally present or associated with "secondary infection." Variola (small-pox), scarlet fever, and measles are excellent examples. It is possible that the danger at the present time is rather in the direction of supposing that every disease will readily yield its secret to the bacteriologist. Such, of course, is not the case. Nevertheless, as in the past, so in the future, constant research and patient investigation is the only hope we have for the elucidation of truth in respect to the causes of disease. About HackerNoon Book Series: We bring you the most important technical, scientific, and insightful public domain books. This book is part of the public domain. Newman, George. 2015.Bacteria. Urbana, Illinois: Project Gutenberg. Retrieved May 2022 from https://www.gutenberg.org/files/48793/48793-h/48793-h.htm This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org , located at https://www.gutenberg.org/policy/license.html .